
Journal of Gastroenterology Research and Practice
Research Article - Open Access, Volume 6
Evaluation of the ability of three HBsAg quantitative test systems to detect variant HBsAg in occult HBV infection
Yang Liu1*; Jianhua Cun2; Chengshan He1; Xiudi Jiang1; Zhicheng Lu1
1Department of Clinical Laboratory, The Seventh People’s Hospital of Shanghai University of Traditional Chinese Medicine,
Shanghai, China.
2Department of Clinical Laboratory, Nujiang Lisu Autonomous Prefecture People’s Hospital, Yunnan, China.
*Corresponding Author : Yang Liu
Department of Clinical Laboratory, The Seventh People’s
Hospital of Shanghai University of Traditional Chinese
Medicine, Shanghai, China.
Email: ly5580246@163.com
Received : Nov 20, 2025
Accepted : Dec 24, 2025
Published : Jan 05, 2026
Archived : www.jjgastro.com
Copyright : © Liu Y (2026).
Abstract
Objective: To analyze the effects of HBV S-region mutations on HBsAg expression, secretion, and antigenicity, and evaluate three clinical HBsAg detection systems.
Methods: Ten high-frequency S-region mutations from OBI patients were selected. Recombinant plasmids were constructed via point mutation, transfected into L02 hepatocytes, and tested after 96h. Cell supernatants and lysates were analyzed using YP, KH, and YHL HBsAg detection systems. Non-reactive samples underwent LC MS/MS for HBsAg confirmation.
Results: Sequencing confirmed correct plasmid construction. V106G mutant HBsAg was undetectable by all three systems but confirmed via LC-MS/MS. YHL missed C85R, K122E, C124R, C138R, and V190A mutants; KH missed C124R, C138R, and V190A; YP missed only C85R, C138R, and V190A in supernatants. Mutations C85R, S114P, C124R, C138R, W172C, and V190A reduced HBsAg secretion, while C85R, L109P, and K122E increased intracellular accumulation.
Conclusion: Mutations variably reduced HBsAg detection, with C85R, C124R, C138R, and V190A showing significant declines. YP demonstrated superior sensitivity, while YHL required improvement. Intracellular accumulation was observed for L109P, K122E, and C85R mutants. Antigenic changes, secretion reduction, and reagent sensitivity contributed to detection variability.
Keywords: Occult Hepatitis B virus infection; Hepatitis B virus surface antigen; Mutation; Quantitative testing
Citation: Yang L, Jianhua C, Chengshan H, Xiudi J, Zhicheng L. Evaluation of the ability of three HBsAg quantitative test systems to detect variant HBsAg in occult HBV infection. J Gastroenterol Res Pract. 2026; 6(1): 1240.
Introduction
Hepatitis B Virus (HBV) infection is a widespread problem worldwide and posesa major threat to human health. Accord ing to the latest data published by the World Health Organisa tion (WHO) in 2020, about 257 million people worldwide have chronic hepatitis B [1]. Each year, approximately 887,000 peo ple die from end-stage liver diseases such as decompensated cirrhosis, liver failure and primary liver cancer caused by HBV infection, and this data clearly reflects that the problem of HBV infection has not been alleviated [2]. Hepatitis B surface anti gen (HBsAg) is an important serological indicator for detecting HBV infection [3]. However, some patients may have the special phenomenon of HBsAg-negative and HBV DNA-positive, which is called Occult Hepatitis B Virus This condition is called Occult Hepatitis B Virus Infection (OBI) [4-6], in which the virus can be transmitted through blood transfusion, organ transplantation, and dialysis, leading to a variety of serious diseases, such as Liv er Cirrhosis (LC) and Hhepatocellular Carcinoma (HCC), which are very dangerous to patients’ health [7]. However, due to the special serological characteristics of OBI, it may lead to prob lems such as underdiagnosis and HBV reactivation during the di agnostic process, a phenomenon that has attracted widespread attention in the medical community. It not only represents a potential health threat to patients, but also poses a challenge to medical treatment strategies and prognostic judgement.
HBsAg is the envelope protein of HBV and is a key target for virus neutralisation by specific HBsAb [8]. Literature reports by analysing HBV gene sequences from patients with OBI have identified multiple amino acid substitutions in HBsAg that are usually missed at the time of testing [9]. Therefore, this variant HBsAg is important for the diagnosis of HBV infection and vac cine development.
Abbott ARCHITECT i2000 Chemiluminescent Immunoas say System, Aphilon iFlash 3000-G Chemiluminescent Immu noassay System, and Kovacs Polaris i2400 Chemiluminescent Immunoassay System are the more widely used systems for quantitative HBsAg detection in clinical practice. We used gene recombination technology to construct eukaryotic expression vectors for the 11 high-frequency mutation sites of the HBV S gene detected from the sera of OBI patients and expressed them in the L02 hepatocyte cell line, and collected cell culture supernatant and cell lysates as samples, and detected the levels of HBsAg in these samples by using the three chemilumines cence immunoassay systems mentioned above, and evaluated the effects of the different assays on the detection of HBsAg concentrations inside and outside the cells. The sensitivity and leakage rate of the different detection systems on the detection of HBsAg concentration inside and outside the cells were evalu ated. Meanwhile, preliminary observation was made to see whether the 10 high-frequency mutation sites of HBV S gene affected the exocytosis of HBsAg.
Materials and methods
Study samples and HBV DNA extraction
After a total of 42 HBsAg-negative (Abbott i2000 chemilu minescent micro particle immunoassay system) serum samples from outpatients and inpatients of the Shanghai Seventh Peo ple’s Hospital with HBV DNA levels ranging from 20 IU/mL to 200 IU/mL (PerkinElmer, USA) and sera of samples from chroni- c hepatitis B patients with a clinical history of more than three years were co- llected in a preliminary stage for the study [10]. In this study, the top 10 mutati- on sites of HBV DNA S-region in the order of frequency in OBI sera, which were not present in the CHB patients in this study, were selected: I81T, C85R, V106G, L109P, S114P, K122E, C124R, C138R, W172C, V190A, and the wild B and C types of HBV as the subjects of this study. This study was approved by the Medical Ethics Committee of our hospital (2018-IRBQYYS-020). DNA was extracted from patient serum using the Serum/Plasma Circulating DNA kit extrac-tion kit (TIANGEN) and frozen at -80°C for subsequent experiments.
HBV DNA PCR amplification and sequencing
HBV S region amplification of extracted DNA was performed using a self -constructed two-round PCR method [11] with primers F: CCTKCTCGTTAGGCGG; R: CGRGCAACGGGGGTAAACG (K and Rare concatenated bases, in which K: G/T and R: A/G), which were synthesised by Huazin Biotechnology. The first and second rounds of PCR were carried out using Premix TapTM polyme- rase to amplify the HBV S region gene. 1% agarose gel electrophoresis (120 V for 30 min) was performed on the am plified products, and the target bands were cut from the gel after electrophoresis, and the DNA gel was recovered using a DNA Gel Recovery Kit (DiaSpin Column PCR Fraction Purification Kit, Shanghai Sangon) for purifying the PCR products. The puri fied positive amplifi cation products were sequenced directly or loaded into a T vector clone, T-Vec tor pMDTM18 (Dalian Baozhi Biotech), and sequenced on a sequencer (ABI37 30, ABI, USA) by Viwit Biotech Co. Sequencing sequences were compared with BLAST of nucleic acid sequences using the NCBI website (https://blast. Ncbi.nlm.nih.gov/Blast.cgi) to determine wheth er the gene sequences were those of the S region of HBV DNA, and the sequences were compared with BLAST of nucleic acid sequences using the website (www.ncbi.nlm.nih.gov /projects/ ge- notyping/) to determine whether the gene sequences of the HBV DNA were those of the S region of HBV DNA. Genotyping/) was used to determine the HBV genotype of each sample, and the nucleic acid and amino acid mutation sequences were ana lyzed using the DNAMAN version 8.08.798 software to compare the reference sequences of each genotype with its counterpart.
Construction of S-region gene mutation plasmid and cell transfection
The wild-type B-type AB602818 and C-type AB014381 HBV DNA S-region genes, which were clearly sequenced without S-region mutation, were selected as templates, and the frag ments containing mutated S-region genes were cloned into pcD NATM3.1/HisA eukaryotic expression vectors (a gift from the Central Laboratory of Changhai Hospital, Shanghai, China) using the PCR targeted mutation (BBI of Shanghai Bio-engineering) and seamless cloning techniques, and transformed into DH5α receptor cells (Suzhou New Saimi Biotechnology) shaking the bacteria coated plate, and sequencing, the sequencing accurate bacterial fluid using endotoxin-free plasmid macroextraction kit (TIANGEN) to extract plasmid, using spectrophotometer to de tect plasmid concentration. The point mutation primers (Table 1) were synthesised by Shanghai Bioengineering.
Table 1: Point mutation primers.
| Primer name | Primer sequence |
|---|---|
| External primers - B-F | TTGGTACCGAGCTCGGCCACCATGCATCATCACCATCACCATGAGAACATCG CATCAGGACTCC |
| External primers - B-R | CTGGATATCTGCAGAATTTTAAATGTATACCCAAAGACAAAAGAAAATT |
| External primers - C-F | TTGGTACCGAGCTCGGCCACCATGCATCATCACCATCACCATGAGAACACAA CATCAGGATTCCTAGGAC |
| External primers - C-R | CTGGATATCTGCAGAATTTCAAATGTATACCCAAAGACAAAAGAAA I81T-FTGTCTGCGGCGTTTTACCATCTTCC |
| I81T-R | GGATGCAGAGGAAGATGGTAAAACGCC |
| C85R-F | TCTGCGGCGTTTTATCATCTTCCT |
| C85R-R | CAGGATGCGGAGGAAGATGATAAAACGC |
| V106G-F | TACCAAGGTATGTTGCCCGGTTGTCC |
| V106G-R | GGCAACATACCTTGGTAGTCCAGAAGAA |
| L109P-F | CGTTTGTCCTCCACTTCCAGGAACATCAACTAC |
| L109P-R | CGTGCTGGTAGTTGATGTTCCTGG |
| S114P-F | GTCCTCTACTTCCAGGAACACCAACTACCAG |
| S114P-R | CCTGGAAGTAGAGGACAAACGGG |
| K122E-F | GCACAACTCCTGCTCAAGGAACCTCTATGT |
| K122E-R | TGAGCAGGAGTTGTGCAGGTCTCGCATGG |
| C124R-F | CAAGACCCGCACAACTCCTGCTCAAGG |
| C124R-R | AGTTGTGCGGGTCTTGCATGGTCC |
| C138R-F | TCTATGTTTCCCTCTTGTTGCTGTACAAAAC |
| C138R-R | GTCCGAAGGTTTTGTACAGCAACAAGA |
| W172C-F | TTTCTCCTGTCTCAGTTTACTAGTGCCATTTGTTCA |
| W172C-R | GAACCACTGAACAAATGGCACTAGT |
| V190A-F | CCACTGCTTGGCTTTCAGTTATATGGATGATGTGG |
| V190A-R | CCCCCAATACCACATCATCCATATAACTG |
L02 cells (source) were inoculated into 6-well plates and pre pared for transfection after the cell density reached 70%~80% as observed under the microscope. 1.5 mL EP tubes were filled with 250 μL of Opti-MEM serum-reduced medium (source), 2500 ng of plasmid, 5μL of Invitrogen Lipofectamine™ 3000 (Thermo Fisher Scientific) and 5μL of P3000 (source) and left at room temperature for 15 min. 250μL of mixed liposomes were added dropwise into the cell wells. ) and 5 μL P3000 (source), and leave it at room temperature for 15 min. 250 μL of the mixed liposomes were added drop by drop into the cell wells, and the six-well plate was shaken gently after adding one well to make the liquid distributed evenly, and the six-well plate was put into 37℃ for incubation after transfection.
Detection of variant HBsAg levels
Cell culture supernatant was collected after 96H of transfec tion; 300 μL of cell lysate was added to each well of cultured cells; 250 μL of lysate was collected, the precipitate was cen trifuged and discarded, and the concentration of protein was adjusted to a quantitative concentration of 0.25 mg/mL with physiological saline.The collected cell culture supernatant and the lysate were divided into three portions, and the lysate was analysed using the ARCHITECT i2000 SR (Abbott, YP) and the supporting The ARCHITECT i2000 SR (Abbott, YP) and accom panying reagents and calibrators; Polaris i2400 (Kehua, KH) and accompanying reagents and calibrators; and iFlash 3000-G (YHL) and accompanying reagents and calibrators; were used for the quantitative detection of HBsAg. The testing process was car ried out in accordance with the instructions of the instruments and reagents, and the relevant indoor quality control products were purchased from Shanghai Clinical Laboratory Centre.
Identification of mutant HBsAg proteins with the V106G mutation in the HBV S region gene
Mutant HBsAg with V106G mutation in HBV S region gene was analysed by using Vanquish Neo/ Orbitrap Exploris 480 liquid chromatography-mass spectrometry (Thermo Fisher Sci entific) with L02 culture supernatant and cell lysate prepared under the same conditions at the same protein concentration. The K122E mutated HBsAg was used as a positive control.
Results
Establishment of a 10-point mutant eukaryotic expression vector for the HBV S gene
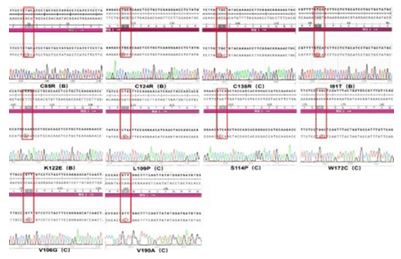
After the I81T/C85R/V106G/L109P/S114P/K122E/C124R/ C138R/W172C/V1 90A point mutant was inserted into the eu karyotic expression vector pcDNA3.1, the sequencing of PCR products were all correct by comparison, as shown in Figure 1.
Ability of three assay systems to detect recombinant mutant HBsAg in cell supernatants and lysates
Three chemiluminescent HBsAg detection systems, YP, KH and YHL, and the accompanying quantitative detection reagents were used to detect the concentration of variant HBsAg in the cell culture supernatant and cell lysate of recombinant plasmid after 96H transfection into L02 cells. The results showed that the three assay systems had 100% positive compliance and nega tive compliance rates for the detection of both empty (negative control) and wild-type samples (positive control). However, the three detection systems were not reactive to the supernatants and cell lysates of cells transfected with mutant locus V106G, resulting in the missed detection of HBsAg. YHL detection sys tem mutant C85R, K122E, C124R, C138R and V190A superna tants and lysates were also not detected. kH detection system mutant C124R and L162R supernatants and lysates were not detected in addition. C85R, C138R and V190A were also not de tected in cell culture supernatants. YP detection system was not detected in cell culture supernatants of cells transfected with C85R, C138R and V190A loci. The specific detection capabilities are shown in Table 1.
Table 1: Results of three assay systems for detection of recombinant mutant HBsAg in cell supernatants and lysates.
|
Sample type Mutation sites/ Detection systems |
Cell culture supernatant (IU/mL) | Cell lysate (IU/mL) | ||||
|---|---|---|---|---|---|---|
| YP | KH | YHL | YP | KH | YHL | |
| NC (pc3.1) | 0.01 | 0.02 | 0.01 | 0.01 | 0.01 | 0.01 |
| wild B type of HBV | 1.57 | 0.4 | 0.79 | 0.19 | 0.12 | 0.29 |
| wild C type of HBV | 1.21 | 0.56 | 0.73 | 0.23 | 0.13 | 0.32 |
| I 81T (B) | 0.83 | 0.39 | 0.37 | 0.16 | 0.11 | 0.19 |
| C85R (B) | 0.01 | 0.02 | 0.01 | 0.57 | 0.07 | 0.01 |
| V106G (C) | 0.01 | 0.01 | 0.01 | 0.04 | 0.02 | 0.02 |
| L109P (C) | 0.2 | 0.13 | 0.29 | 0.47 | 0.06 | 0.15 |
| S114P (C) | 1.2 | 0.25 | 0.59 | 0.15 | 0.11 | 0.16 |
| K122E (B) | 1.13 | 0.12 | 0.01 | 0.35 | 0.11 | 0.03 |
| C124R (B) | 0.06 | 0.03 | 0.01 | 0.09 | 0.05 | 0.01 |
| C138R (C) | 0.01 | 0.01 | 0.01 | 0.07 | 0.08 | 0.01 |
| V172C (C) | 0.98 | 0.37 | 0.68 | 0.15 | 0.09 | 0.17 |
| V190A (C) | 0.01 | 0.02 | 0.01 | 0.19 | 0.05 | 0.03 |
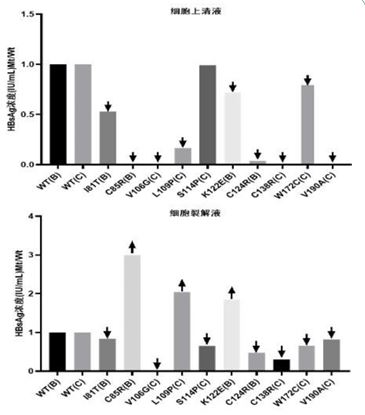
Differences in detection of variant HBsAg levels by the three assay systems
In order to compare the differences in sensitivity and con tent of the three assay systems for detecting mutant HBsAg, the HBsAg concentration in the culture supernatant and cell lysate of HBV wild B-type and C-type transfected cells for 96 hr from each assay system was used as the target value, and the ratios of the HBsAg concentration in the culture and cell lysate of the transfected cells at the 10 S-region mutated loci to the target value were calculated (mutated/wild, Mt/Wt) ) (Table 2).
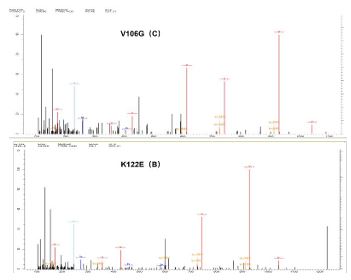
Identification of variant HBsAg with the V106G mutation in the S region of HBV
In this study three clinically used HBsAg quantitative de tection systems did not detect variant HBsAg with the V106G mutation. To verify the presence of HBsAg variant proteins in these samples, we subjected the variant HBsAg with the V106G mutation to LC-MS/MS for protein identification analysis, and the variant HBsAg with the K122E mutation served as a positive control. The results of protein identification analysis were as follows (Figure 3). Mutant HBsAg was present in both samples sent for testing.
Discussion
HBsAg encoded by HBV S region is an important serological biomarker for HBV detection, which can generally be detected by kits 4-10 weeks after HBV infection, so HBsAg has an irre placeable role in screening early HBV infection, and it can also be used to evaluate the therapeutic efficacy of the patient and to judge the progression of the disease [12]. However, with the development of detection technology, we found that a part of the patients whose HBsAg is undetectable using current tech nology but whose HBV DNA is positively expressed, and such patients are referred to as OBI [13]. The 2017 European Associa tion for the Study of the Liver (EASL) Clinical Guidelines on Hep atitis B Virus Infection divide the natural history of HBV infec tion into five stages [14], of which the fifth stage refers to OBI. The characteristic of OBI is the persistent presence of HBV DNA in liver tissue or serum, which cannot be detected using exist ing diagnostic reagents (excluding the window period for HBV infection). Obviously, OBI is not only a potential threat to the safety of the blood supply, but also has the potential for reacti vation of HBV when the patient develops an immunodeficiency or undergoes immunosuppressive therapy, which may result in acute exacerbation of hepatitis, cirrhosis, and hepatocellular carcinoma [15]. In patients with OBI or in some chronic hepati tis B patients, mutations, insertions and deletions in the HBV S region genes can cause immunogenic changes in HBsAg, reduce or prevent replication of viral particles, and inhibit the secretory function of HBsAg [16]. These may all be factors contributing to OBI, making HBsAg not well detected by the commercial HBsAg testing systems commonly used in clinical practice today.
HBsAg, an outer membrane protein of HBV, was first pro posed by BLUMBERG [17] in 1965, and is the most clinically used indicator for determining HBV infection, and qualitative and quantitative assays have been widely used in blood screen ing, HBV infection, and in the clinical management and efficacy of medications used for chronic hepatitis B. In order to investi gate the role of HBV S-region specific amino acid substitutions in HBV OBI-infected patients on the expression, secretion, and antigenicity of variant HBsAg, and thus the consequent impact on the detectability of commercially available HBsAg detec tion systems commonly used in the clinic, we constructed the I81T, C85R, V106G,L109P, S114P, K122E, C124R, C138R, W172C, V190A 10 S-region gene mutation eukaryotic expression plas mids, respectively, transfected with human liver cell line L02, and the culture supernatant (secretory function) and cell lysate (cell accumulation) were taken to simulate the clinical samples, to evaluate the detection ability of YP, KH, and YHL assay sys tems on HBsAg quantitative detection system. The results showed that the mutant HBsAg produced by mutation V106G was missed in all three detection systems, both in cell culture supernatants and cell lysates, but protein identification analysis by LC-MS/MS confirmed the presence of mutant HBsAg in the samples. We believe that this result may be due to the signifi cant change in the antigenicity of mutant HBsAg produced by this mutation site, which makes the capture antibody/detection antibody of the detection kit unable to recognise mutant HBsAg well, resulting in missed detection. capture antibody/detection antibody does not recognise the variant HBsAg well, resulting in missed detection. Compared with the corresponding HBV wild-type HBsAg levels, the levels of variant HBsAg in both cell culture and cell lysate samples were reduced to different de grees, among which C85R, C124R, C138R and V190A showed a significant reduction and even missed detection. This indicates that the variant HBsAg expressed by these 10 high-frequency mutation sites in the S region obtained from OBI-infected pa tients can affect the ability of the detection system to detect HBsAg, which may be related to the change in the antigenicity of the variant HBsAg, the decrease in the secretion of the vari ant HBsAg, and the sensitivity of the detection reagents. Among the three detection systems, the YP detection system was the most prominent in the ability to identify variant HBsAg and de tection sensitivity, which was significantly better than the other two detection systems; the YHL detection system needs to be further improved and perfected in the ability to identify variant HBsAg, and it failed to detect variant HBsAg generated by the C85R, K122E, C124R, C138R and V190A mutations. The results of the study showed that the detection of mutated HBsAg.
The results of the 10 S-region mutation loci in the YP assay system were used to calculate the ratio of the concentration of variant HBsAg to the concentration of wild-type HBsAg in the two sample types, and it was found that the concentration of wild-type HBsAg were significantly higher in the culture super natant than in the cell lysates, which were 8.26 and 5.26 times, respectively, and the three assay systems behaved in this way. The concentrations of the variant HBsAg produced by the muta tions of 10 S-region mutation loci were relatively similar in the cell culture and cell lysates. The concentrations of HBsAg in cell culture fluid and cell lysate were relatively similar, even the con centrations of variant HBsAg cell lysate produced by mutations at the L109P, K122E and C85R loci were significantly higher than those in cell culture fluid, 12.39-fold, 2.92-fold, and 3-fold, re spectively. The results suggest that the mutant HBsAg produced by the S-region mutation sites in this paper all have some de gree of impaired secretion function, especially the mutant HB sAg produced by the mutations at the L109P, K122E and C85R sites showed intracellular accumulation. This phenomenon can seriously affect the analytical performance of the commercial HBsAg detection system, resulting in missed detections; at the same time, the accumulation of variant HBsAg in hepatocytes may induce the development of endoplasmic reticulum stress, which is one of the causative factors of hepatocellular carcino ma.
Of course, there are still many shortcomings in this study, the study is limited to cross-sectional collection of samples, limited to the S region of the HBV genome, the future needs to estab lish a cohort study with a larger sample size, long-term and ef fective monitoring of the whole genome of HBV changes in the process of the development of OBI patients. The mutation of 10 loci in the S region of HBV found in this study can have a large impact on the three quantitative HBsAg detection systems com monly used in the clinic, resulting in a certain degree of missed detection. It is hoped that the domestic detection system will further improve the analytical performance and variant identifi cation to meet the clinical needs.
References
- Li H, Yan L, Shi Y, et al. Hepatitis B virus infection: overview. Adv Exp Med Biol. 2020; 1179: 1–16.
- Lindner P, Christensen SB, Nissen P, et al. Cell death induced by the ER stressor thapsigargin involves death receptor 5, a non-autophagic function of MAP1LC3B, and distinct contributions from unfolded protein response components. Cell Commun Signal. 2020; 18: 12.
- Liu H, Chen S, Liu X, et al. Effect of S-region mutations on HBsAg in HBsAg-negative HBV-infected patients. Virol J. 2024; 21: 92.
- Raimondo G, Locarnini S, Pollicino T, et al. Update of the statements on biology and clinical impact of occult hepatitis B virus infection. J Hepatol. 2019; 71: 397–408.
- Wang H, Wang M, Huang J, et al. Novel hepatitis B virus surface antigen mutations associated with occult genotype B hepatitis B virus infection affect HBsAg detection. J Viral Hepat. 2020; 27: 915–921.
- Saitta C, Pollicino T, Raimondo G. Occult hepatitis B virus infection: an update. Viruses. 2022; 14: 1504.
- Mak LY, Wong DK, Pollicino T, et al. Occult hepatitis B infection and hepatocellular carcinoma: epidemiology, virology, hepatocarcinogenesis and clinical significance. J Hepatol. 2020; 73: 952–964.
- D’Souza S, Lau KC, Coffin CS, et al. Molecular mechanisms of viral hepatitis-induced hepatocellular carcinoma. World J Gastroenterol. 2020; 26: 5759–5783.
- Jiang X, Chang L, Yan Y, et al. Role of S protein transmembrane domain mutations in the development of occult hepatitis B virus infection. Emerg Microbes Infect. 2022; 11: 2184–2196.
- He C, Liu Y, Jiang X, et al. Frequency of HBsAg variants in occult hepatitis B virus-infected patients and detection by ARCHITECT HBsAg quantitative assay. Front Cell Infect Microbiol. 2024; 14: 1368473.
- He C, Jiang X, Xu Z, et al. Amplification of low-load HBV DNA S region genes using a self-developed two-round PCR method and optimisation of experimental conditions. J Second Mil Med Univ. 2021; 42: 1157–1163.
- Tseng TC, Liu CJ, Yang H, et al. High levels of hepatitis B surface antigen increase risk of hepatocellular carcinoma in patients with low HBV load. Gastroenterology. 2012; 142: 1140–e14.
- Ikeda K, Kobayashi M, Someya T, et al. Occult hepatitis B virus infection increases hepatocellular carcinogenesis eightfold in patients with non-B, non-C liver cirrhosis: a cohort study. J Viral Hepat. 2009; 16: 437–443.
- Liu T, Liu A, Liu Y, et al. In vitro investigation of HBV clinical isolates from Chinese patients reveals that genotype C isolates possess higher infectivity than genotype B isolates. Virol Sin. 2022; 37: 398–407.
- Hsu CW, Yeh CT. Emergence of hepatitis B virus S gene mutants in patients experiencing hepatitis B surface antigen seroconversion after peginterferon therapy. Hepatology. 2011; 54: 101–108.
- Blumberg BS, Sutnick AI, London WT. Australia antigen and hepatitis. JAMA. 1969; 207: 1895–1896.