
Journal of Gastroenterology Research and Practice
Review Article - Open Access, Volume 5
Function of the esophageal mechanoreceptors
Ivan M Lang, DVM, PhD*
Dysphagia Research Laboratory, Division of Gastroenterology and Hepatology, Department of Medicine, Medical College of Wisconsin, USA.
*Corresponding Author : Ivan M Lang
Dysphagia Research Laboratory, Division of Gastroenterology and Hepatology, Department of Medicine, Medical College of Wisconsin, Milwaukee, WI 53225, USA.
Tel: 414 456-8138;
Email: imlang@mcw.edu
Received : Jan 10, 2025
Accepted : Feb 04, 2025
Published : Feb 11, 2025
Archived : www.jjgastro.com
Copyright : © Lang IM (2025).
Abstract
There are four histologically defined esophageal mechanoreceptive agents with evidence-based functions, three are mechanoreceptors and one is a mechanoreceptive inhibitory afferent nerve. The slowly-adapting tension sensitive receptors of the esophageal myenteric plexus function to prevent Esophago-Pharyngeal Reflux (EPR) through the Esophago-Upper Esophageal Sphincter Contractile Reflex (EUCR), Esophago-Esophageal Contractile Reflex (EECR), and Secondary Peristalsis (SP). The rapidly-adapting touch sensitive receptors of the esophageal mucosa function to promote EPR and airway protection during belching through esophago-upper Esophago-Upper esophageal sphincter Relaxation Reflex (EURR), Esophago-Glottal Closure Reflex (EGCR), Esophago-Hyoid Distraction Reflex (EHDR), and Esophageal Retrograde Peristaltic Reflex (ERPR). The slowly-adapting tension receptors of the mucosa function to promote bolus transport from pharynx to stomach by activating the Esophago-Pharyngeal Swallow Reflex (EPSR) instead of SP, especially in infant humans and animals, and ensuring that the esophageal phase of swallowing follows the pharyngeal phase. The afferent receptive inhibitory motor neurons innervating the LES and crural diaphragm function to eliminate the esophago-gastric barrier, i.e., LES and crural diaphragm contraction, during swallowing and belching. There are also undefined mechanoreceptors of the esophagus which activate splanchnic nerve afferents and probably mediate autonomic and nociceptive functions. However, the specific receptors as well as responses involved have not been identified.
Keywords: Esophagus; Mechanoreceptors; Mucosa; Muscularis.
Citation: Lang IM. Function of the esophageal mechanoreceptors. J Gastroenterol Res Pract. 2025; 5(1): 1223.
Introduction
There have been many review articles of the anatomy, histology and sensory nature of the esophageal mechanoreceptors, but none have also fully reviewed the functions of these receptors [1-6]. There were three basic types of studies used to define the esophageal mechanoreceptors and their functions: a) histological studies, which identified and localized possible mechanoreceptors [7-16], b) physiological studies which defined the afferent neural responses of mechanical stimulation of the esophageal wall [15-21], and c) physiological studies which defined the reflex effects of mechanical stimulation of the esophageal wall [6,10,22-49]. While these studies identified and characterized the esophageal mechanoreceptors or identified the neural or physiological effects of various esophageal stimuli, there were none that had defined esophageal mechanoreceptors and their functions in the same experiments. Therefore, for this review histological studies were analyzed first to identify the known mechanoreceptors, and the other studies were used to apply the appropriate reflex responses and functions to each mechanoreceptor.
Esophageal mechanoreceptors
Slowly adapting muscular mechanoreceptors
The first esophageal mechanoreceptor observed using histological techniques [7,12,13] was the receptor of the myenteric plexus which was described as an Intraganglionic Laminar Ending (IGLE [12,13]). Independent structures within the ganglia, which have spindle shaped thickenings towards their endings, are mostly found in the upper and lower portions of the esophagus and make contact with the vagus nerve [7]. These characteristics suggested that these structures are sensory in function and probably mechanoreceptive [7,12,13]. Later studies [16] found that either contraction or distension of the esophagus activated the IGLE’s of the myenteric plexus, thereby defining them as mechanoreceptors.
The first step to identifying the function of this mechanoreceptor was defining its response characteristics and sensory innervation. It was found that applying tension to the esophageal wall by inflating a balloon intraluminally activated vagal afferents and did this in a slowly-adapting manner [8,11,16-22]. That is, the rate of vagal response was directly related to the magnitude of the tension applied to the esophagus and did not stop responding until the tension stopped. Other types of stimuli, like stroking the mucosa, had no effect on this receptor [8,19,20]. Therefore, this myenteric plexus mechanoreceptor was innervated by vagal afferents and operated in slowly-adapting manner.
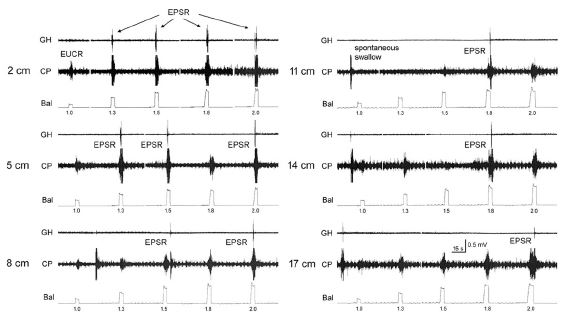
It was found that distension or contraction of the esophagus [24,29,32,35] was associated with contraction of the esophagus orad of the stimulus (Esophago-Esophageal Contractile Reflex (EECR)) and/or the upper esophageal sphincter (Esophago-UES Contractile Reflex (EUCR)), and usually followed by Secondary Peristalsis (SP). The most significant study [30] associating the slowly-adapting muscular mechanoreceptor with these reflexes was the observation that removal of the mucosal layer of the esophagus blocked all reflexes stimulated by distension of the esophagus except the EUCR, EECR, and SP (Figure 1). Considering that no other receptor had been found in the muscular layer or that anesthetizing the mucosa with lidocaine did not block SP or EUCR [30], these reflexes must have been mediated by muscular mechanoreceptors. This was further confirmed by the observations that the EUCR and SP were more readily activated by a slowly rather than rapidly applied tension to the esophageal wall and that these reflexes were not activated by stroking the esophageal mucosa [30]. Therefore, the slowly-adapting muscular mechanoreceptor mediates EUCR, EECR, and SP.
The functions of EUCR, EECR and 2P are complimentary, and when acting together unified. The EECR acts to block or inhibit the orad movement of any esophageal bolus orad of the stimulus [35], the EUCR acts to prevent esophago-pharyngeal reflux of the bolus [30] and SP acts to move distally a bolus orad of the stimulus [3,6]. Therefore, since all three reflexes are activated by the same stimulus in the same region of the esophagus [30], these three reflexes have a unified function of preventing any bolus remaining in the esophagus after the passage of peristalsis to be refluxed to the pharynx.
Rapidly adapting mucosal mechanoreceptors
The first study suggesting that there was a receptor in the mucosa of the esophagus found intraepithelial fibers lining the wall within the mucosa [14]. This location and structure suggested that they had a sensory function. It was later found that this mucosal receptor is a vagally mediated mechanoreceptor [17,18], but the specific nature of the stimulus activating this receptor was identified later. In later more controlled studies [8,19,20], it was found that the vagally mediated mechanoreceptors of the esophageal mucosa are best stimulated by stroking the mucosa, rather than applying tension to the esophageal wall. In addition, it was found that these mechanoreceptors are rapidly-adapting [8,19,20], that is, the primary response is at the beginning or ending of the stimulus, and they respond best to a rapidly applied stimulus.
The function of these rapidly-adapting mucosal mechanoreceptors was defined by experiments which investigated the effects of removal of the mucosa from the esophagus. It was found that removal of this layer of the esophagus totally eliminated the activation of belching [30] and the reflex effects that occur during belching, but had no effect on EUCR, EECR or SP (Figure 1). These belch-related reflexes [30,34,50] are esophago-upper esophageal sphincter (UES) relaxation reflex (EURR) (Figures 1, 3, 4 & 6), Esophago-Glottal Closure Reflex (EGCR) (Figures 1 & 2), Esophago-Hyoid Distraction Reflex (EHDR) (Figures 1 & 3), and the Esophageal striated muscle Retrograde Peristaltic Response (ERPR) (Figure 4). Belching and all of these associated reflexes are activated by rapid injection of air into the esophagus [30,34,50], which correlates very well with the rapidly-adapting nature of the mucosal mechanoreceptors. In addition, belching and all of these reflexes are mediated by the vagus nerve [30,34], which is the afferent nerve of the rapidly-adapting mucosal mechanoreceptors. Therefore, the rapidly adapting mucosal mechanoreceptors mediate belching and its associated reflexes, which aid the process: EURR, EGCR, EHDR and ERPR.
Slowly adapting tension/mucosal mechanoreceptors
In one early study [15] esophageal mechanoreceptors were identified by recording responses from vagal afferent fibers innervating the cervical esophagus during esophageal contraction or respiration. One set of receptors acted like the slowly-adapting muscular tension receptors, but a second set activated by esophageal distension exhibited responses which had characteristics of both slowly-adapting and rapidly-adapting mechanoreceptors. That is, at the beginning of the distension the receptor was maximally activated and this response declined even though the stimulus did not, similar to a rapidly-adapting receptor. The response of the receptor then acted like a slowly-adapting receptor as the rate of the response was directly related to the strength of the distension [15]. In later more detailed studies [20], it was found that there are slowly-adapting tension mechanoreceptors in the mucosa, which also respond to touch [20].
The function of the mucosal slowly-adapting tension mechanoreceptors has not been defined, but there are two studies which strongly suggest a function. Stimulation of the Esophagus can activate the Pharyngeal Swallow (EPSR) (Figure 5), but this response is more sensitive in animals [33] and infant humans [51,52] than adult humans [59]. In cat studies [33], it was found that the EPSR is activated in a probabilistic manner. That is, the larger the bolus, the longer area of the esophagus stimulated, and the closer the stimulus is to the UES, the more likely it is for the stimulus to activate the pharyngeal swallow. The EPSR [33] was blocked by lidocaine applied to the esophageal mucosa, and intraluminal injection of HCl into the esophagus increased the probability of activation of EPSR [33]. These studies suggested that the EPSR is activated by stimulation of receptors in the mucosa of the esophagus. Considering that the EPSR is not secondary peristalsis, secondary peristalsis is mediated by muscular mechanoreceptors [30], the EPSR is activated by tension to the esophageal wall [33], and EPSR [33] but not SP [53], is blocked by luminal application of lidocaine, it was concluded that the EPSR is mediated by the slowly-adapting tension mechanoreceptors of the esophageal mucosa [33].
The physiological function of the EPSR was hypothesized to be a mechanism which guaranteed bolus transport during swallowing, thereby, preventing aspiration [33,54]. The EPSR is most sensitive in animals [33,54] and infant humans [51,52], and in both the larynx is high in the neck [55]. This high position allows the epiglottis to go behind the soft palate and the larynx to open directly into the nasopharynx [56]. If a swallowed bolus gets stuck in the upper esophagus and secondary peristalsis is activated, this contraction of the upper esophagus could push the bolus orad as well as caudad and cause esophago-pharyngeal reflux and aspiration. However, if instead of activating secondary peristalsis, the pharyngeal swallow is activated this response would push the bolus down the esophagus without any possibility of reflux or aspiration [56]. This scenario of esophageal reflux in human infants is not unreasonable or unlikely as esophageal reflux induced aspiration is the primary theory accounting for sudden infant death syndrome [54,57,58].
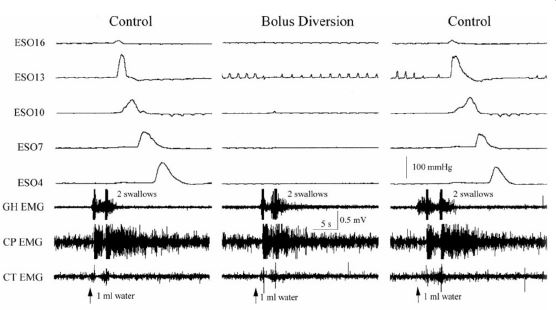
Another way in which the mucosal slowly-adapting tension mechanoreceptors function to promote bolus transport is by ensuring that the esophageal phase of swallowing follows the pharyngeal phase. It has been observed in various species [30,60,61] that diversion of the swallowed bolus from the esophagus prevents the initiation of the esophageal phase of swallowing (Figure 6). This occurs not only for swallows with a bolus, but also for non-bolus swallows [30]. Given that the receptor of this function must be very sensitive and close to the mucosal surface, it is highly likely that activation of the mucosal slowly adapting tension receptors mediates this function. Therefore, the mucosal slowly adapting tension mechanoreceptors of the esophagus activate two reflex responses that function to promote the transmission of the swallowed bolus from pharynx to stomach and prevent aspiration.
Mechanosensitive esophageal inhibitory motor innervation
There is one more receptive process of the esophagus that alters physiologic function, but it is not through a separate unique receptor, it is through inhibitory motor nerves with receptor properties [10,23,25,29,36,37]. Unlike the esophageal mechanoreceptors described above, the function of this receptive process was identified and characterized before the receptive process [10,23,25,29,36,37].
The first indication of this receptor was in the studies which found that stretching or contraction of the distal esophagus caused the Lower Esophageal Sphincter (LES [23,25] and crural diaphragm [36,37] to relax. A later study [29] of the mechanism of this reflex inhibition of the LES found that this response was not blocked by vagotomy, sympathectomy, hexamethonium or atropine, but was blocked by tetrodotoxin. This study determined that this response was neurally mediated, but not mediated by a peripheral or central reflex [29]. Therefore, this effect must have been due to mechanical activation of the inhibitory motor nerve [29]. It was later determined that the motor inhibitory neurons from the esophagus to the LES and crural diaphragm have a mechanoreceptive function [10].
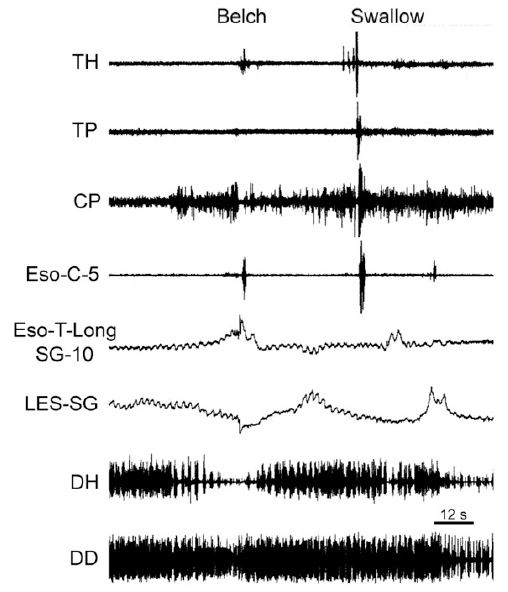
The function of this mechanosensitive inhibitory innervation is elimination of the esophago-gastric barrier, i.e., LES and crural diaphragm contraction. This response is an integral part of both swallowing and belching. During swallowing the peristaltic process activates distal esophageal longitudinal contraction [23,37,42,63] which is closely associated with both LES [23,29,62] and crural diaphragm [36,37,62] relaxation. Similarly, during belching (Figure 7) the gastric air bolus reflexly activates distal esophageal longitudinal contraction before initiation of the belch, and this response is closely associated with relaxation of both the LES and crural diaphragm [29,39,40,49,50]. The actual belch does not occur until the esophago-gastric barrier is reduced enough to allow gastro-esophageal reflux of the air bolus [50,62] which activates the rapidly adapting mucosal receptors [30,50], as described above.
Sympathetic mechanoreceptors
It has been found that distension of the esophagus activates two types of receptors that activate splanchnic nerve afferents [9]. One type is a slowly-adapting muscular tension receptor and the other is a rapidly-adapting touch sensitive serosal receptor.
The function of these receptors is unknown, but since they activate sympathetic afferents, they probably affect the autonomic nervous system. Mechanical stimulation of the esophagus can increase blood pressure [35,41,46] and cause pain [41,64]. Therefore, these esophageal sympathetic activating mechanoreceptors may provide receptive function for these responses. Further studies are needed to identify the mechanoreceptors, and define their responses and functions.
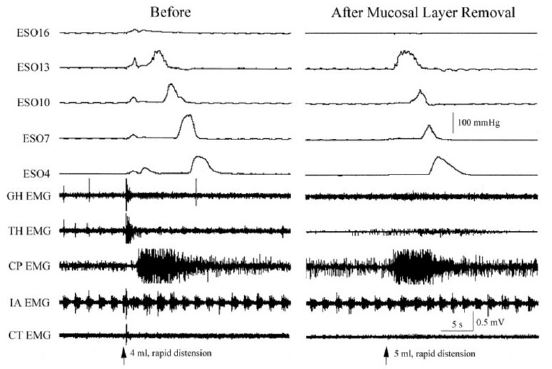
ESO#: Esophagus #; LES: Lower Esophageal Sphincter; EMG: Electromyography; HG: Geniohyoideus; TH: Thyrohyoideus; CP: Cricopharyngeus; IA: Intraaryternoideus; CT: Cricothyroideus.
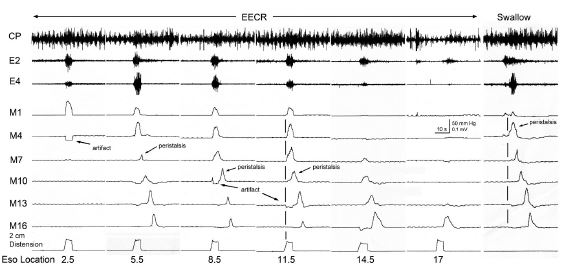
CP: Cricopharyngeus; SP: Secondary Peristalsis; UES: Upper Esophageal Sphincter; E#: EMG Recording # cm from the CP; M#: Manometry Recording # cm from CP; Eso location: Location of the balloon from the CP.
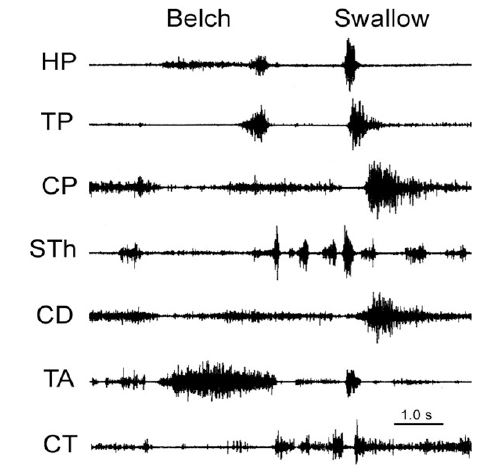
EGCR: Esophago-Glottal Closure Reflex; HP: Hypopharyngeus; TP: Thyropharyngeus; CP: Cricopharyngeus; STh: Stylohyoideus; CD: Cricoarytenoideus Dorsalis; TA: Thyroarytenoideus; CT: Cricothyroideus.
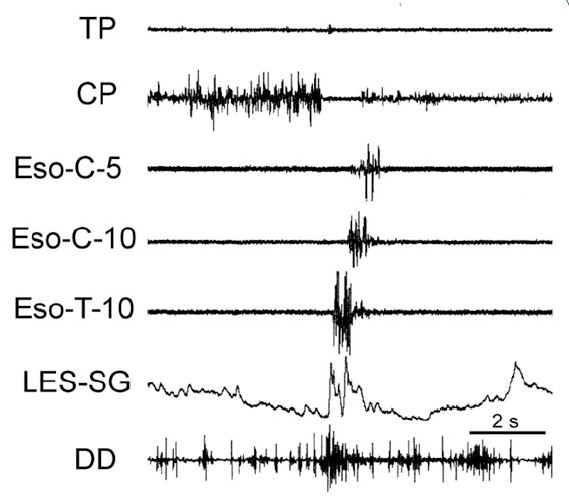
LES: Lower Esophageal Sphincter; EURR: Esophago-UES Relaxation Reflex; TP: Thyropharyngeus; CP: Cricopharyngeus; Eso-C-#: Cervical esophagus # cm from the CP; Eso-T-#: Thoracic esophageuas #cm from the LES.
GH: Geniohyoideus; CP: Cricopharyngeus; Bal: Ballon Tension.
ESO#: Esophageal Manometry #cm from the LES; GH: Geniohyoideus; CP: Cricopharyngeus; CT: Cricothyroideus; EMG: Electromyography.
TH: Thyrohyoideus; TP: Thyropharyngeus; CP: Cricopharyngeus; Eso: Esophagus; C: Cervical; T: Thoracic; LES: Lower Esophageal Sphincter; DH: Diaphragmatic Hiatus; DD: Diaphragmatic Dome; SG: Strain Gauge.
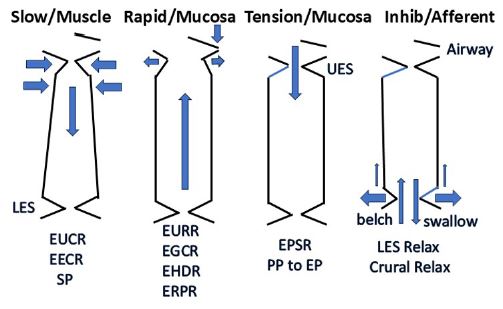
UES: Upper Esophageal Sphincter; LES: Lower Esophageal Sphincter; EUCR: Esophago-UES Contractile Reflex; EECR: Esophago-Esophageal Contractile Reflex; SP: Secondary Peristalsis; EURR: Esophago-UES Relaxation Reflex; EGCR: Esophago-Glottal Closure Reflex; EHDR: Esophago-Hyoid Distraction Reflex; ERPR: Esophago-Retrograde Contraction Reflex; EPSR: Esophago-Pharyngeal Swallow Reflex; PP: Pharyngeal Phase; EP: Esophageal Phase.
Conclusion
There are three identified esophageal mechanoreceptors with defined functions. The slowly-adapting muscular mechanoreceptors mediate the following reflex functions which prevent esophago-pharyngeal reflux: EUCR, EECR and SP. The rapidly-adapting mucosal mechanoreceptors mediate belching and its associated reflex responses: EURR, EGCR, EHDR, and ERPR. The slowly-adapting mucosaltension receptors function to promote bolus transport from pharynx to stomach by activating the EPSR instead of SP, especially in infant humans and animals, and ensuring that the esophageal phase of swallowing follows the pharyngeal phase. In addition, there is the mechanoreceptive inhibitory motor innervation of the LES and crural diaphragm which facilitates emptying of the esophagus during swallowing and gastro-esophageal reflux during belching. Undefined esophageal receptors mediate mechanoreceptive sympathetic afferent activation which probably have nociceptive functions.
Acknowledgements: The support and financial assistance of Dr. Reza Shaker and technical assistance of Dr. Bidyut Medda were greatly appreciated.
References
- Christensen J. Origin of sensation in the esophagus. Am J Physiol. 1984; 246: G221–G225.
- Diamant NE. Physiology of esophageal motor function. Gastroenterol Clin NA. 1989; 18: 179–200.
- Goyal RK, Chaudbury A. Physiology of normal esophageal motility. J Clin Gastroenterol 2008; 42: 610–619.
- Lang IM. The Physiology of Esophageal Reflexes. OBM Hepatology and Gastroenterology. 2020; 4: 35.
- Paintal AS. Vagalsensory receptors and their reflex effects. Physiol Rev. 1973; 53: 159–210.
- Schoeman MN, Holloway RH. Stimulation and characteristics of secondary oesophageal peristalsis in normal subjects. Gut. 1994; 35: 152-158.
- Asaad K, Rahman AE, Nawar NNY, Mikhail Y. Intrinsic innervation of the oesophagus with special reference to the presence of muscle spindles. Acta Anat. 1983; 115: 91-96.
- Clerc N, Mei N. Vagal mechanoreceptors located in the lower oesophageal sphincter of the cat. J Physiol. 1983; 336: 487–498.
- Clerc N, Mei N. Thoracic esophageal mechanoreceptors connected with fibers following sympathetic pathways. Brain Res Bull. 1983; 10: 1–7.
- Dong H, Jiang Y, Dong J, Mital RK. Inhibitory motor neurons of the esophageal myenteric plexus are mechanosensitive. Am J Physiol. 2015; 308: C405–C413.
- Falempin M, Mei N, Rousseau JP. Vagal mechanoreceptors of the inferior thoracic oesophagus, lower oesophageal sphincter and the stomach in the sheep. Pfulgers Archiv. 1978; 373: 25–30.
- Nonidez JF. Afferent endings in the ganglia of the intermuscular plexus of the dog’s esophagus. J Comp Neurol. 1946; 85: 177–189.
- Rodrigo J, Hernandez J, Vidal MA, Pedrosa JA. Vegetative innervation of the esophagus. II. Intraganglionic laminar endings. Acta Anat. 1975; 92: 79-100.
- Rodrigo J, Hernandez J, Vidal MA, Pedrosa JA. Vegetative innervation of the esophagus. III. Intraepithelial endings. Acta Anat. 1975; 92: 242–258.
- Satchell PM. Canine oesophageal mechanoreceptors. J Physiol. 1984; 346: 287-300.
- Zagorodnyuk VP, Brookes SJH. Transduction sites of vagal mechanoreceptors in the guinea pig esophagus. J Neurosci. 2000; 20: 6249–6255.
- Mei N. Etude electrophysiologique des recepteurssensibles de l’oesophagethoracique du Chat. CR Acad Sc. 1965; 260: 302–305.
- Mei N. Mecanorecepteursvagaux digestifs chez le chat. Exp Brain Res. 1970; 11: 502–514.
- Page AJ, Martin CM, Blackshaw LE. Vagal mechanoreceptors and chemoreceptors in mouse stomach and esophagus. J Neurophysiol. 2002; 87: 2095–2103.
- Page AJ, Blackshaw LA. An in vitro study of the properties of vagal afferent fibres innervating the ferret oesophagus and stomach. J Physiol. 1998; 512: 907–916.
- Sekizawa SI, Ishikawa T, Sant’Ambroglio FB, Sant’Ambroglio G. Vagal esophageal receptors in anesthetized dogs: mechanical and chemical responsiveness. J Appl Physiol. 1999; 867: 1231–1235.
- Andrew BL. The nervous control of the cervical oesophagus of the rat during swallowing. J Physiol. 1956; 134: 729–740.
- Babaei A, Bhargava V, Korsapati H, Zheng WH, Mittal RK. A unique longitudinal muscle contraction associated with transient lower esophageal sphincter relaxation. Gstroentenrology. 2008; 134: 1322-331.
- Craemer B, Schlegel J. Motor responses of the esophagus to distension. J Appl Physiol. 1957; 10: 498 – 504.
- Dogan I, Bhargava V, Liu J, Mittal RK. Axial stretch: a novel mechanism of the lower esophageal sphincter relaxation. Am J Physiol. 2007; 292: G329–G334.
- Franzi SJ, Martin CJ, Cox MR, Dent J. Response of canine lower esophageal sphincter to gastric distension. Am J Physiol. 1990; 259: G380–G385.
- Goyal RK, Rattan S. Nature of the vagal inhibitory innervation to the lower esophageal sphincter. J Clin Invest. 1975; 55: 119–126.
- Heywood LY, Wood AK. Retrograde oesophageal contractions in the dog. QJ Exp Physiol. 1988; 73: 87-94.
- Jiang Y, Bhargava V, Mittal RK. Mechanism of stretch-activated excitation and inhibition responses in the lower esophageal sphincter. Am J Physiol. 2009; 297: G397–G405.
- Lang IM, BK Medda, R Shaker. Mechanisms of reflexes induced by esophageal distension. Am J Physiol. 2001; 281: G1246-G1263.
- Lang IM, Medda BK, Jadcherla S, R Shaker. The role of the superior laryngeal nerve in esophageal reflexes. Am J Physiol. 2012; 302: G1445-G1457.
- Lang, IM, BK Medda, A Babaei, R Shaker. The role of peripheral reflexes in the initiation of the esophageal phase of swallowing. Am J Physiol. 2014; 306: G728-G737.
- Lang IM, Medda BK, Jadcherla SR. Shaker R. Characterization and mechanisms of the pharyngeal swallow activated by stimulation of the esophagus. Am J Physiol. 2016; 311: G827-G837.
- Lang IM, Medda BK, Shaker R. Characterization and mechanisms of the supragastric belch in the cat. Am J Physiol. 2017; 313: G220-G229.
- Lang. IM, Medda, BK, Shaker R. Characterization and mechanisms of the esophago-esophageal contractile reflex of the striated muscle esophagus. Am J Physiol. 2019; 317: G304–G313.
- Liu J, Yamamoto Y, Schirmer BD, Ross RA, Mittal RK. Evidence for a peripheral mechanism of esophago-crural diaphragm inhibitory reflex in cats. Am J Physiol. 2000; 278: G281–G288.
- Liu J, Puckett JL, Takeda T, Jung HY, Mitttal RK. Crural diaphragm inhibition during esophageal distension correlates with contraction of the esophageal longitudinal muscles in cats. Am J Physiol. 1993; 288: G927–G932.
- Loomis CW, Yao D, Bieger D. Characterization of an esophagocardiovascular reflex in the rat. Am J Physiol. 1997; 272: 783–791.
- McNally EF, Kelly JE, Ingelfinger FJ. Mechanisms of belching: effects of gastric distension with air. Gastroenterology. 1964; 64: 254–259.
- Monges H, Salducci J, Naudy B. Dissociation between the electrical activity of the diaphragmatic dome and crura muscular fibers during esophageal distension, vomiting, and eructation. An electromyographic study in the dog. J Physiol (Paris). 1978; 74: 541–554.
- Pickering M, Campion D, Jones JFX. Reflex cardiorespiratory effects of nociceptive oesophageal distension in the decerebrate rat. Exp Physiol. 2002; 87: 41–48.
- Pouderoux P, Lin S, Kahrilas PJ. Timing, propagation, coordination, and effect of esophageal shortening during peristalsis. Gastroenterology. 1997; 112: 1147–1154.
- Oliven A, Haxiu M, Kelsen SG. Reflex effect of esophageal distension on respiratory muscle activity and pressure. J Appl Physiol. 1989; 66: 536–541.
- Paterson WG. Neuromuscular mechanisms of esophageal responses at and proximal to a distending ball. Am J Physiol. 1991; 260: G148–G155.
- Paterson WG, Ratan S, Goyal RK. Esophageal responses to transient and sustained esophageal distension. Am J Physiol. 1988; 255: G587–G595.
- Satpathy NK, Al-Sattar NA. The effects of acute oesophageal distension on arterial blood pressure, E.C.G. and respiration in dog. Ind J Physiol Pharm. 1984; 28: 105–114.
- Shaker R, Ren J, Kern M, Dodds WJ, Hogan WJ, Li Q. Mechanisms of airway protection and upper esophageal sphincter opening during belching. Am J Physiol. 1992; 262: G621–G628.
- R Shaker, J Ren, B Medda, IM Lang, V Cowles, S Jaradeh. Identification and characterization of the esophago-glottal closure reflex in cat. Am J Physiol. 1994; 266: G147 -G153.
- Straahof JW, Ringers JK, Lamers CB, Masclee AA. Provocation of transient lower esophageal sphincter relaxation by gastric distension with air. Am J Gastroenterology. 2001; 96: 2317–2323.
- Lang, IM, BK Medda, R Shaker. The digestive and respiratory tract motor responses associated with eructation. Am J Physiol. 2013; 304: G1044-G1053.
- Jadcherla SR, Duing HQ, Hoffman RG, Shaker R. Esophageal body and upper esophageal sphincter motor responses to esophageal provocation in preterm newborns. J Pediatr. 2003;143: 31–38.
- Jadcherla SR, Hoffman RG, Shaker R. Effect of maturation of the magnitude of mechanosensitive and chemosensitive reflexes in the premature human esophagus. J Pediatr. 2006; 149: 77–82.
- Chen CL, Lu TT, Yi CH. Effects of lidocaine on esophageal secondary peristalsis. Neurogastroenterol Motil. 2010; 22: 606–610.
- Lang IM, Medda BK, Shaker R, Jadcherla S. The effect of body position on esophageal reflexes in cats: A possible mechanism of SIDS?. Pediatr Res. 2018; 83: 731-738.
- Laitman JT, Reidenberg JS. Specialization of the human respiratory and upper digestive system as seen through comparative and developmental anatomy. Dysphagia. 1993; 8: 318–25.
- Bremner RM, Hoeft SF, Costantini M, Crookes PF, Bremner CG, Demeester TR. Pharyngeal swallowing. The major factor in clearance of esophageal reflux episodes. Ann Surgery. 1993; 218: 364–370.
- Al-Adnani M, Cohen MC, Scheinberg I. Gastroesophageal reflux disease and sudden infant death syndrome: Mechanisms behind an underrecognized association. Pediatr Dev Pathol. 2011; 14: 53–56.
- Page M, Jeffrey H. The role of gastro-esophageal reflux in the etiology of SIDS. Early Human Dev. 2000; 59: 127–149.
- Taniguchi H, Aoyagi Y, Matsuo K, Imaeda S, Hirumuta M, Saitoh E. Elicitation of the swallowing reflex by esophageal stimulation in healthy subjects. Dysphagia. 2020; 35: 657–666.
- Longhi EH, Jordan PH. Necessity of a bolus for propagation of primary peristalsis in the canine esophagus. Am J Physiol. 1971; 220: 609–612.
- Janssens J, Valembois P, Hellemans J. Studies on the necessity of a bolus for the progression of secondary peristalsis in the canine esophagus. Gastroenterology. 1974; 67: 245–252.
- Lang IM. The physiology of eructation. Dysphagia. 2016; 31: 121–133.
- Mittal RK, Padda B, Bhalla V, Bhargava V, Liu J. Synchrony between circular and longitudinal muscle contractions during peristalsis in normal subjects. Am J Physiol2006; 290: G431 – G438.
- Drewes AM, Schipper KP, Dimcevski G, Petersen P, Andersen OK, Gregersen H, Arenst-Nielsen L. Multimodal assessment of pain in the oesophagus: a new experimental model. Am J Physiol2002; 283: G95 – G103.