
Journal of Gastroenterology Research and Practice
Review Article - Open Access, Volume 4
Exploring the interaction between bile acid and intestinal microbiota in acute-on-chronic liver failure and the progress of treatment strategies
Yuyu Zeng1,2; Dakai Gan2; Yuhe Zeng3; Molong Xiong1,2*
1The Third Affiliated Hospital, Jiangxi Medical College, Nanchang University, China.
2Infectious Diseases Hospital Affiliated to Nanchang University, China.
3Shanxi Medical University, China.
*Corresponding Author : Molong Xiong
Infectious Diseases Hospital Affiliated to Nanchang
University, Nanchang 330000, No.167, Qingshan Lake
Avenue, Nanchang City, China.
Tel: 139-7094-9572;
Email: xml86005@163.com
Received : Mar 23, 2024
Accepted : Apr 22, 2024
Published : Apr 29, 2024
Archived : www.jjgastro.com
Copyright : © Xiong M (2024).
Abstract
Acute-on-Chronic Liver Failure (ACLF) is an acute and severe disease involving single or multiple organ failure. Due to the unclear pathogenesis, there is still a lack of effective drug treatment. Intestinal microbiota is closely related to Bile Acid (BA) metabolism. In the progression of chronic liver disease to ACLF, BA metabolism disorder is often accompanied by intestinal microbiota imbalance, which affect one another mutually. This article reviewed the role of BA metabolism and intestinal microbiota in the progression of ACLF, which aims to explore potential new targets for the treatment of ACLF.
Keywords: Bile acid; Intestinal microbiota; Acute-on-chronic liver failure; Pathophysiology; Treatment strategy.
Citation: Zeng Y, Gan D, Zeng Y, Xiong M. Exploring the interaction between bile acid and intestinal microbiota in acute-onchronic liver failure and the progress of treatment strategies. J Gastroenterol Res Pract. 2024; 4(4): 1195.
Introduction
Acute-on-Chronic Liver Failure (ACLF) is a severe liver failure that occurs in the presence of chronic hepatitis or cirrhosis, and it is a highly aggressive and challenging medical condition [1]. Despite the significant advancements in Artificial Liver Support System (ALSS) in recent years and the introduction of different combined ALSS for treating ACLF, the mortality rate at 28-day and 90-day remain alarmingly high, reaching 33% and 58% respectively [2]. Due to the significant disease burden associated with ACLF, it is crucial to elucidate pathogenesis and explore novel treatment strategies.
The liver and the intestine are closely connected, with 70% of the liver’s blood supply originating from the portal vein, which gathers nearly all the blood from the intestine, indicating that intestinal products may play an important role in the progression of ACLF. Although the pathogenesis of ACLF is not fully understood, some scholars believe that systemic inflammatory plays a crucial role in the pathophysiological mechanisms of ACLF [3]. Partial patients with cirrhosis release a large number of Pathogen-Associated Molecular Patterns (PAMPs) and Damage-Associated Molecular Patterns (DAMPs), which activate innate immune cells through binding to their specific Pattern Recognition Receptors (PRRs), resulting in excessive systemic inflammatory response, due to immune-mediated serious tissue damage, such as liver, kidney, lung or other organs appear successively or simultaneously failure, and eventually progressing ACLF [4-6].
The liver serves as the body’s metabolic hub, which synthesize and metabolize Bile Acid (BA). BA function as versatile signaling molecules, regulating the communication between liver and intestine [7,8]. The BA pool in normal humans slightly fluctuates daily within a certain range [9]. When ACLF occurs, the synthesis and metabolism of BA is disturbed and the intestinal microbiota is dysregulated. These alterations contribute to further liver injury [10-12]. This review aims to (a) provide an overview of the toxic effects of BAs on hepatocytes; (b) discuss the alterations in the intestinal microbiota of ACLF; (c) provide an overview of the connections between BA, intestinal microbiota and ACLF; and (d) elucidate the potential therapeutic role of targeting BA metabolism and intestinal microbiota in treating ACLF.
Metabolism of BA and their toxic effects on hepatocytes
As shown in Figure 1, the liver synthesizes primary BA, such as Cholic Acid (CA) and Chenodeoxycholic Acid (CDCA) from cholesterol [13]. This can generally occur through two synthetic pathways: classical and alternative pathways. The classical pathway produces CA and CDCA through the enzymatic action of CYP7A1, CYP8B1 and CYP27A1 cholesterol hydroxylases. The alternative pathway is the synthesis of CDCA through the hydroxylation of cholesterol side chains by CYP27A1, followed by the 7α-hydroxylation of 27-hydroxycholesterol and other oxysterols by CYP7B1 [14]. After synthesis, primary BA bind with glycine or taurine to create primary conjugated BA. These are stored in the gallbladder along with bile. When cholecystokinin is activated, typically after a meal, they are eventually released into the intestinal lumen. When the terminal ileum is reached, the intestinal epithelium’s surface reabsorbs 95% of the BAs in the form of glycine or taurine conjugated BA through the Apical Sodium-Dependent Bile Acid Transporter (ASBT, also called IBAT). These, along with the absorbed substances, enter the portal vein. Hepatocytes uptake BAs from the blood of portal circulation through the Na+-Taurocholate Contra sporting Polypeptide (NTCP) and Organic Anion Transporting Polypeptide (OATP), and then excrete them into the intestine via the biliary system. This process is called the enterohepatic circulation of BA [15]. Additionally, a small portion of primary BA reaches the colon intact, where specific intestinal microbiota converts it into secondary BA, which contains Deoxycholic Acid (DCA) and Lithocholic Acid (LCA). These secondary BA are either passively reabsorbed by the colonic mucosa or excreted in the feces, constituting around 5% of the BA pool [16].
BAs are amphiphilic molecules containing hydrophilic hydroxyl group, carboxyl group and hydrophobic alkyl group. The hydrophobicity is inversely related to the number of hydroxyl group in the side chain, and the order of hydrophobicity BAs is as follows: LCA>DCA>CDCA>CA [17]. The toxicity of BAs is closely related to their hydrophobicity, the greater the hydrophobicity, the less soluble in water at the same concentration, and the stronger the toxicity.
Cholestasis is a pathological state in which bile flow cannot enter the duodenum and enter the blood instead due to the obstruction of bile formation, secretion and excretion caused by various reasons [18]. The pathological changes of ACLF are one-time (or multiple) necrosis of hepatocytes, which can be manifested as mass necrosis (the necrosis scope exceeds 2/3 of the hepatic parenchyma), and sub-mass necrosis (about 1/2 ~ 2/3 of the hepatic parenchyma) [19]. The residual hepatocytes are regenerated to different degrees, and the fine and small bile duct hyperplasia, cholestasis and bile embolism can be observed. These changes in the structure of the bile duct, especially the capillary bile duct and fine bile duct partial disappearance, compression, distortion, etc., aggravated cholestasis [10].
Maintaining BA concentration below a specific threshold is crucial to prevent hepatotoxicity. Early in vitro experiments have shown that when toxic BA is added to hepatocytes, the submillimolar level of toxic BA directly damage hepatocytes [20,21]. Through studies on primary human hepatocytes, Woolbright et al. [22] demonstrated that the primary cause of human cholestatic hepatitis is the leakage of BA from the biliary system back into the liver parenchyma, resulting in direct hepatocytes necrosis. However, the levels of toxic BA in the serum and tissue levels rarely reach these submillimolar levels in patients with ACLF, which suggests that BA may cause liver damage in other ways.
Li et al. demonstrated that Taurocholic Acid (TCA) can stimulate the expression of inflammatory cytokines, and their study highlights the importance of cytokines in initiating BA-induced inflammatory responses [23]. Similarly, Cai et al. found significantly increased cytokines expression level by applying pathophysiological concentrations of Glycochenodeoxycholic Acid (GCDCA) in human hepatocyte medium. These increased cytokines are mainly the chemokine family, including CCL15, CCL20, CXCL1, etc. [24]. These experiments all show that when BAs accumulate in hepatocytes, they can cause liver damage by triggering an inflammatory response [24].
In addition, Cai et al. showed that neutrophils play an important role in patients with cholestatic hepatitis, suggesting that cholestatic hepatitis is mediated by inflammatory neutrophilic response and its associated mitochondrial damage and oxidative stress, while Kupffer Cells (KCs) seem to be unrelated to this damaging response [24]. However, other researchers have different opinion, suggesting that KCs are involved in the BA induced liver damage response. KCs are activated after Bile Duct Ligation (BDL) and causes BA accumulation by affecting the function of BA transporters [25]. The use of gadolinium chloride (an inhibitor of KCs) in the BDL model mitigated liver damage [26], suggesting that KCs play an important role in promoting BDL-induced liver damage. Which may be related to the activation of NF-kB and JNK signaling pathways.
The imbalance of intestinal microbiota in ACLF patients exacerbates disease progression
The intestinal microbiota, also known as the microbial community, is a varied assemblage of microorganisms found within the human digestive system. The intestinal microbiota of one person is typically formed by the age of one and remains stable for the rest of their life, with occasional minor changes. In adults, the intestinal microbiota comprises around 1014 microorganisms, which is almost tenfold greater than the number of human cells. This microbiota encompasses 4-10 primary phylas and a range of 3000-5000 species [27]. The dominant bacteria in the human intestinal microbiota are from four main groups (phyla) - Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria. These groups make up over 97% of the total microbiota. The adult intestinal microbiota varies greatly among individuals and is primarily influenced by factors such as host genetics, diet, and the environment. The intestinal microbiota has a significant impact on the metabolic processes, immune regulation, development, and physiology of the host organism [28].
Over the past few years, numerous research studies have commenced exploring the correlation between the variety and makeup of the human intestinal microbiota and ACLF. Some related researches revealed that alterations in the intestinal or serum microbiota of ACLF patients (Table 1) [12,29-32]. However, there are some differences between these studies, which may be due to the etiology of liver disease, sequencing methods, diet, environment, and individual differences [29,31]. In addition, results obtained using cutting-edge research techniques suggest that changes in composition of intestinal microbiota can affect liver disease progression [33].
Imbalance in the intestinal microbiota can contribute to chronic liver disease, and multiple researches have demonstrated that changes in the intestinal microbiota have a significant impact on the development and advancement of liver disease. As chronic liver disease progresses, patients experience changes in intestinal neurological function, resulting in damage to the intestinal barrier and reduced secretion of antimicrobial peptides, which leads to intestinal inflammation [34,35]. Additionally, there is an impaired barrier and increased intestinal permeability, allowing bacteria and components from the intestine to be transported to the liver through the portal vein [36,37]. This compromises the liver’s immune system by producing PAMPs and Microbial-Associated Molecular Patterns (MAMPs), causing or worsening the systemic inflammatory response and ultimately exacerbating the progression of ACLF [38,39].
Interaction between BAs and intestinal microbiota in ACLF patients
The intestinal microbiota participates in the anabolism of BAs, which enriches the diversity of BA pool. First, the activity of acyl-CoA synthetase required for the conversion of unconjugated BA to conjugated BA is regulated by the intestinal microbiota [40]. Second, when the primary conjugated BA enters the intestine, it is uncoupled by Bile Salt Hydrolase (BSH), and the glycine or taurine groups are removed [41]. Song et al found that microbiota with BSH activity including Staphylococcus, Bacillus, Paenibacillus, Clostridium, and Brevibacillus [42]. Finally, in the intestine, the primary BA is converted to secondary BA by the action of 7α-dehydroxylase [43]. The bacteria with dehydroxylation activity were distributed in Bacteroides, Escherichia, Clostridium and Lactobacillus [44]. In addition, the intestinal microbiota can also modify BA molecule through different isomerization, which is one of the main mechanisms for increasing BA diversity. For example, Clostridium mediates the 7α/ β-isomerization of CDCA in humans to form Ursodeoxycholic Acid (UDCA) [8].
BA can regulate the composition of intestinal microbiota through the following mechanisms. Firstly, BA can play an antibacterial role by regulating the pH of the intestine and destroying the cell membrane of harmful microbiota [41]. Secondly, high concentration of BA can directly dissolve the bacterial cell membrane, thereby leading to the outflow of enzymes in the cells and hindering bacterial overgrowth [45]. For example, DCA can inhibit the growth of Lactobacillus and Bifidobacterium, while CA can reduce the activity of Roseburia, Lactobacillus, and Ruminococcus [46]. Finally, a phase I study of oral BA erivative Obeticholic Acid (OCA) in healthy people found that oral OCA inhibited the overgrowth of gram-positive bacteria by activating Farnesoid X Receptor (FXR) [47].
To summarize, as shown in Figure 2, there is a two-way relationship between the intestinal microbiota and BA. The intestinal microbiota controls the production of BA by enzymes, while BA influence the makeup of the intestinal microbiota by regulating the pH of the intestine, toxicity or activating FXR.
Novel therapeutic approaches developed based on BA metabolism and the intestinal microbiota Inhibition of the enterohepatic circulation of BA
Inhibition of enterohepatic circulation of BA is a promising treatment for ACLF patients with significantly increased serum BA concentration. ASBT inhibitor can reduce the accumulation of BAs in the liver by inhibiting the reabsorption of BAs by the terminal ileal epithelial cells. A study showed that inhibition of ASBT gene expression could prevent severe cholestatic liver injury in bile duct ligation mice by inhibiting the reabsorption of BAs and increasing bile salt excretion from the kidney[48]. In addition, inhibiting the uptake of BAs by hepatocytes is also a promising therapeutic approach. NTCP plays a key role in the uptake of BAs by hepatocytes, and NTCP inhibitors can reduce the accumulation of BAs in hepatocytes and protect hepatocytes from BAs overload [49]. However, because the specific structure of NTCP has not been clarified, so far, no effective NTCP inhibitors have been developed.
Fecal microbiota transplantation
Fecal Microbiota Transplantation (FMT) involves transferring the complete collection of microorganisms found in the feces of a healthy donor into the recipient’s intestinal in order to restore or modify the makeup and operation of the intestinal microbiota [50]. After antibiotic treatment, FMT has proven to be an effective solution for refractory and recurrent Clostridium Difficile Infection (CDI) [51,52]. Numerous studies have demonstrated that FMT restores the normal structure of fecal bacterial community and fecal BA in patients with CDI, while also increasing the ratio of secondary BA to primary BA [53]. Additionally, FMT has shown promising results in treating Hepatic Encephalopathy (HE) [54,55]. In a randomized controlled trial conducted by Bajaj et al. [56], FMT not only enhanced cognitive function in HE but also increased the diversity of intestinal microbiota and abundance of beneficial bacteria in HE patients.
Additionally, FMT has been utilized for the management of liver failure. A study demonstrated that FMT derived from donors in good health enhanced the survival rate of individuals suffering from Severe Alcohol-Associated Hepatitis (SAH) while also diminishing SAH-related complications [57]. A different clinical trial examined the effectiveness of FMT for alcohol-related ACLF. The trial revealed that the group FMT had considerably higher survival rates at both the 28-day and 90-day marks compared to the group standard medical treatment (100% versus 60% and 100% versus 57.14% respectively). These findings ultimately suggest that FMT is a secure option for alcohol-related ACLF. The utilization of FMT has the potential to enhance the survival rate of patients in the medium term and ameliorate the clinical severity score [58].
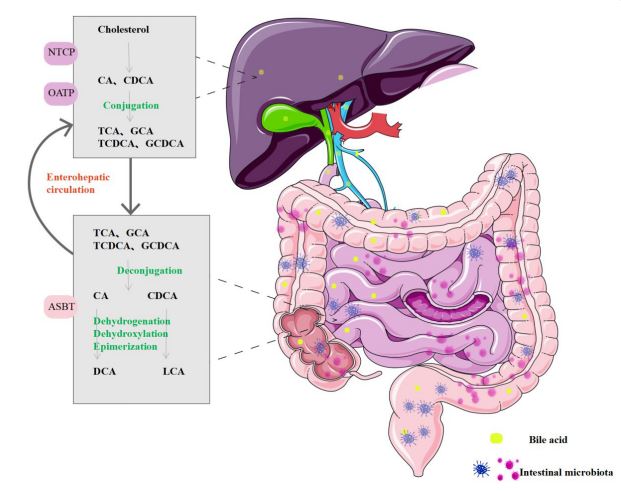
Schematic overview of the enterohepatic circulation: Primary BA is synthesized from cholesterol in the liver and combined with glycine or taurine to form primary conjugated BA, which are excreted into the intestinal lumen with bile. Ninety-five percent of the primary conjugated BA is reabsorbed by enterocytes under the effect of ASBT in the terminal ileum, reflux to the liver through the portal vein, and enter the liver through NTCP/OATP transport. Then, BA pass through the bile ducts into the intestine again, and this process is known as the enterohepatic circulation of BA. The remaining primary BA form secondary BA in response to the intestinal microbiota, which are then passively reabsorbed by the colonic mucosa or excreted in feces.
BA: Bile Acid; ASBT: Apical Sodium-Dependent Bile Acid Transporter; NTCP: Na+-Taurocholate Cotransporting Polypeptide; OATP: Organic Anion Transporting Polypeptide.
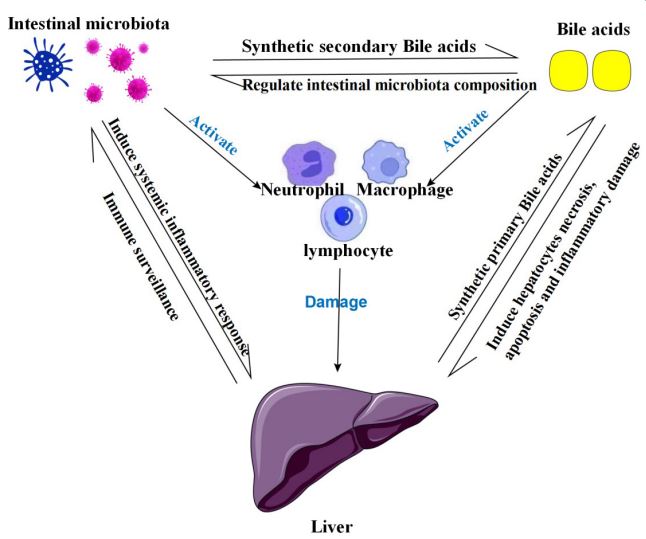
Summary of the liver-BA-intestinal microbiota relationship diagram: The relationship between BA, intestinal microbiota, and liver is not unidirectional, in fact, all these factors are interrelated and interact through different mechanisms. This relationship provides exciting new insights into the pathogenesis, treatment, progression and prognosis of ACLF. In addition, BA and intestinal microbiota can also activate inflammatory cells such as neutrophils, macrophages and lymphocytes to release inflammatory factors such as IL-1, IL-6, IL-8, IL-10, IFN-γ, and TNF-α, which further cause liver injury. BA: bile acid.
Table 1: Changes in intestinal microbiota of ACLF patients in different studies.
| Researchers | Study population | Control population | Sample | Sequencing method |
Increased species abundance |
Decreased species abundance |
|---|---|---|---|---|---|---|
|
Chen et al. 2015 [29] |
79 ACLF patients | 50 healthy controls | Stool | 16S rDNA sequencing | Lachnospiraceae |
Bacteroidaceae,
Ruminococca- ceae, and Lanchnospiraceae |
|
Zhang et al. 2019[30] |
50 HBV-ACLF patients | 23 healthy controls | Plasma |
16S rDNA pyrosequencing |
Moraxellaceae,
Sulfurovum, Comamonas and Burkholderiaceae |
Actinobacteria,
Deinococcus- Thermus, Alphaproteobacte- ria, Xanthomonadaceae and Enterobacteriaceae |
|
Yao et al. 2021 [31] |
91 HBV-ACLF patients | 30 healthy controls | Stool | 16S rDNA sequencing |
Veilonella,
Streptococcus, Enterococcus, and Klebsiella |
Bacteroidetes |
|
Solé et al. 2021 [32] |
65 ACLF patients |
84 decompensated cirrhosis without ACLF |
Stool | Ion Proton Sequencer |
Enterococcus and Pepto- streptococcus |
Roseburia and Firmicutes |
| Wang al. 2021 [12] |
212 ACLF patients |
877 healthy individuals |
Stool | 16S rRNA sequencing | The ratio of cocci to bacilli | Not mentioned |
| Wang al. 2021 [12] |
165 ACLF patients of improved outcomes at discharge |
47 ACLF patients of disease progression at discharge |
Stool | 16S rRNA sequencing | Faecalibacterium | Enterococcus |
Discussion and outlook
With the development of the molecular basis of the relationship between BA homeostasis and intestinal microbiota in the enterohepatic circulation, not only have many novel therapeutic targets for ACLF emerged, but also researchers have begun to study the progression and short-term prognosis of ACLF by BA and intestinal microbiota. According to pertinent research, serum total BA have been found to be associated with the progression of cirrhosis to ACLF [59]. Nevertheless, the predictive significance of total BA and individual BA components in the prognosis of patients with ACLF, whether in the short or long term, remains inadequately researched. Two additional research studies on the properties of the intestinal microbiota in ACLF have indicated that intestinal microbiota displayed varying compositions during different phases of ACLF. The progression of ACLF was linked to a significant presence of Enterococcus, whereas the improvement in ACLF was linked to a substantial presence of Faecalibacterium [12,29]. This result suggests that microbiota characteristics also have potential as prognostic markers in ACLF. However, there are still few studies on this aspect, so future research in this area can be increased. In summary, with the deepening of research, the role of BA-intestinal microbiota will enable us to have a better understanding of the pathogenesis, prognosis and disease management of ACLF.
Abbreviations
ACLF: Acute-on-Chronic Liver Failure; ALSS: Artificial Liver Support System; BA: Bile Acid; CA: Cholic Acid; CDCA: Chenodeoxycholic Acid; ASBT: Apical Sodium-Dependent Bile Acid Transporter; NTCP: Na+-Taurocholate Cotransporting Polypeptide; OATP: Organic Anion Transporting Polypeptide; DCA: Deoxycholic Acid; LCA: Lithocholic Acid; FXR: Farnesoid X Receptor; FGF15/19: Fibroblast Growth Factor 15/19; TGR5: Takeda-GProtein‒Receptor-5; GCDCA: Glycochenodeoxycholic Acid; BDL: Bile Duct-Ligated; CHB: Chronic Hepatitis B; HBV: Hepatitis B Virus; LPS: Lipopolysaccharide; Pamps: Pathogen-Associated Molecular Patterns; Mamps: Microbial-Associated Molecular Patterns; BSH: Bile Salt Hydrolase; UDCA: Ursodeoxycholic Acid; OCA: Obeticholic Acid; FMT: Fecal Microbiota Transplantation; CDI: Clostridium Difficile Infection; IBAT: Ileal Bile Acid Transporter; HE: Hepatic Encephalopathy; RCT: Randomized Controlled Trial; SAH: Severe Alcohol-Associated Hepatitis; SBP: Spontaneous Bacterial Peritonitis.
Declaration of competing interest: The authors have declared that no competing interest exists.
Acknowledgements: This work was supported by Science and Technology Project of Health Commission of Jiangxi Province in 2024 (202410568).
References
- Moreau R, Gao B, Papp M, Bañares R, Kamath PS. Acute-onchronic liver failure: A distinct clinical syndrome. J Hepatol. 2021; 75(1): S27-s35.
- Mezzano G, Juanola A, Cardenas A, Mezey E, Hamilton JP, et al. Global burden of disease: acute-on-chronic liver failure, a systematic review and meta-analysis. 2022; 71: 148-155.
- Arroyo V, Moreau R, Jalan R. Acute-on-Chronic Liver Failure. The New England journal of medicine. 2020; 382: 2137-2145.
- Clària J, Arroyo V, Moreau R. Roles of systemic inflammatory and metabolic responses in the pathophysiology of acute-on-chronic liver failure. JHEP reports: Innovation in hepatology. 2023; 5: 100807.
- Engelmann C, Zhang IW, Clària J. Mechanisms of immunity in acutely decompensated cirrhosis and acute-on-chronic liver failure. Liver Int. 2023.
- Wang S, Zhu H, Pan L, Zhang M, Wan X, et al. Systemic inflammatory regulators and risk of acute-on-chronic liver failure: A bidirectional mendelian-randomization study. Front Cell Dev Biol. 2023; 11: 1125233.
- De Aguiar Vallim TQ, Tarling EJ, Edwards PA. Pleiotropic roles of bile acids in metabolism. Cell Metab. 2013; 17: 657-669.
- Wahlström A, Sayin SI, Marschall HU, Bäckhed F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell metabolism. 2016; 24: 41-50.
- Saran C, Brouwer KLR. Hepatic Bile Acid Transporters and Drug-induced Hepatotoxicity. Toxicologic pathology. 2023; 926233231212255.
- Bao Z, Wei R, Zheng X, Zhang T, Bi Y, et al. Landscapes of gut microbiome and bile acid signatures and their interaction in HBVassociated acute-on-chronic liver failure. Frontiers in microbiology. 2023; 14: 1185993.
- Philips CA, Ahamed R, Abduljaleel JKP, Rajesh S, Augustine P. Identification and Analysis of Gut Microbiota and Functional Metabolism in Decompensated Cirrhosis with Infection. J Clin Transl Hepatol. 2023; 11: 15-25.
- Wang K, Zhang Z, Mo ZS, Yang XH, Lin BL, et al. Gut microbiota as prognosis markers for patients with HBV-related acute-onchronic liver failure. Gut microbes. 2021; 13: 1-15.
- Xiang T, Deng Z, Yang C, Tan J, Dou C, et al. Bile acid metabolism regulatory network orchestrates bone homeostasis. Pharmacol Res. 2023; 196: 106943.
- Jia W, Xie G, Jia W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nature reviews. Gastroenterology & hepatology. 2018; 15: 111-128.
- Perino A, Demagny H, Velazquez-Villegas L, Schoonjans K. Molecular Physiology of Bile Acid Signaling in Health, Disease, and Aging. Physiol Rev. 2021; 101: 683-731.
- Ashby K, Navarro Almario EE, Tong W, Borlak J, Mehta R, et al. Review article: Therapeutic bile acids and the risks for hepatotoxicity. Alimentary pharmacology & therapeutics. 2018; 47: 1623-1638.
- Perez MJ, Briz O. Bile-acid-induced cell injury and protection. World journal of gastroenterology. 2009; 15: 1677-1689.
- Chen J, Zhang S. The Role of Inflammation in Cholestatic Liver Injury. J Inflamm Res. 2023; 16: 4527-4540.
- Li H, Xia Q, Zeng B, Li ST, Liu H, et al. Submassive hepatic necrosis distinguishes HBV-associated acute on chronic liver failure from cirrhotic patients with acute decompensation. J Hepatol. 2015; 63: 50-59.
- Schölmerich J, Becher MS, Schmidt K, Schubert R, Kremer B, et al. Influence of hydroxylation and conjugation of bile salts on their membrane-damaging properties--studies on isolated hepatocytes and lipid membrane vesicles. Hepatology. 1984; 4: 661-666.
- Attili AF, Angelico M, Cantafora A, Alvaro D, Capocaccia L. Bile acid-induced liver toxicity: relation to the hydrophobic-hydrophilic balance of bile acids. Med Hypotheses. 1986; 19: 57-69.
- Woolbright BL, Dorko K, Antoine DJ, Clarke JI, Gholami P, et al. Bile acid-induced necrosis in primary human hepatocytes and in patients with obstructive cholestasis. Toxicology and applied pharmacology. 2015; 283: 168-177.
- Li M, Cai SY, Boyer JL. Mechanisms of bile acid mediated inflammation in the liver. Mol Aspects Med. 2017; 56: 45-53.
- Cai SY, Ouyang X, Chen Y, Soroka CJ, Wang J, et al. Bile acids initiate cholestatic liver injury by triggering a hepatocyte-specific inflammatory response. JCI Insight. 2017; 2: 0e90780.
- Kim SY, Jeong JM, Kim SJ, Seo W, Kim MH, et al. Pro-inflammatory hepatic macrophages generate ROS through NADPH oxidase 2 via endocytosis of monomeric TLR4-MD2 complex. Nat Commun. 2017; 8: 2247.
- Zandieh A, Payabvash S, Pasalar P, Morteza A, Zandieh B, et al. Gadolinium chloride, a Kupffer cell inhibitor, attenuates hepatic injury in a rat model of chronic cholestasis. Hum Exp Toxicol. 2011; 30: 1804-1810.
- He P, Shen X, Guo S. Intestinal flora and linear growth in children. Front Pediatr. 2023; 11: 1252035.
- Zhang X, Luo X, Tian L, Yue P, Li M, et al. The gut microbiome dysbiosis and regulation by fecal microbiota transplantation: umbrella review. Front Microbiol. 2023; 14: 1286429.
- Chen Y, Guo J, Qian G, Fang D, Shi D, et al. Gut dysbiosis in acuteon-chronic liver failure and its predictive value for mortality. Journal of gastroenterology and hepatology. 2015; 30: 1429-1437.
- Zhang Y, Zhao R, Shi D, Sun S, Ren H, et al. Characterization of the circulating microbiome in acute-on-chronic liver failure associated with hepatitis B. Liver international: Official journal of the International Association for the Study of the Liver. 2019; 39: 1207-1216.
- Yao X, Yu H, Fan G, Xiang H, Long L, et al. Impact of the Gut Microbiome on the Progression of Hepatitis B Virus Related Acuteon-Chronic Liver Failure. Frontiers in cellular and infection microbiology. 2021; 11: 573923.
- Solé C, Guilly S, Da Silva K, Llopis M, Le-Chatelier E, et al. Alterations in Gut Microbiome in Cirrhosis as Assessed by Quantitative Metagenomics: Relationship With Acute-on-Chronic Liver Failure and Prognosis. Gastroenterology. 2021; 160: 206-218.e213
- Hsu CL, Schnabl B. The gut-liver axis and gut microbiota in health and liver disease. Nature reviews. Microbiology. 2023; 21: 719-733.
- Wiest R, Albillos A, Trauner M, Bajaj JS, Jalan R. Targeting the gut-liver axis in liver disease. J Hepatol. 2017; 67: 1084-1103.
- Wang L, Fouts DE, Stärkel P, Hartmann P, Chen P, Llorente C et al. (2016) Intestinal REG3 Lectins Protect against Alcoholic Steatohepatitis by Reducing Mucosa-Associated Microbiota and Preventing Bacterial Translocation. Cell Host Microbe. 19: 227-239
- Albillos A, De Gottardi A, Rescigno M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J Hepatol. 2020; 72: 558-577
- Goode EC, Warburton RC, Gelson WT, Watson AJ. Activated intestinal macrophages in patients with cirrhosis release NO and IL-6 that may disrupt intestinal barrier function. Gastroenterology. 2013; 145: 1481-1484.
- Trebicka J, Bork P, Krag A, Arumugam M. Utilizing the gut microbiome in decompensated cirrhosis and acute-on-chronic liver failure. Nature reviews. Gastroenterology & hepatology. 2021; 18: 167-180.
- Tripathi A, Debelius J, Brenner DA, Karin M, Loomba R, et al. The gut-liver axis and the intersection with the microbiome. Nat Rev Gastroenterol Hepatol. 2018; 15: 397-411.
- Sayin SI, Wahlström A, Felin J, Jäntti S, Marschall HU, et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 2013; 17: 225-235.
- Ridlon JM, Harris SC, Bhowmik S, Kang DJ, Hylemon PB. Consequences of bile salt biotransformations by intestinal bacteria. Gut microbes. 2016; 7: 22-39.
- Song Z, Cai Y, Lao X, Wang X, Lin X, et al. Taxonomic profiling and populational patterns of bacterial bile salt hydrolase (BSH) genes based on worldwide human gut microbiome. Microbiome. 2019; 7: 9.
- Ridlon JM, Daniel SL, Gaskins HR. The Hylemon-Björkhem pathway of bile acid 7-dehydroxylation: history, biochemistry, and microbiology. J Lipid Res. 2023; 64: 100392.
- Funabashi M, Grove TL, Wang M, Varma Y, Mcfadden ME, et al. A metabolic pathway for bile acid dehydroxylation by the gut microbiome. Nature. 2020; 582: 566-570.
- He S, Li L, Yao Y, Su J, Lei S, Zhang Y et al. Bile acid and its bidirectional interactions with gut microbiota: a review. Crit Rev Microbiol. 2023; 1-18.
- Zhan K, Zheng H, Li J, Wu H, Qin S, et al. Gut Microbiota-Bile Acid Crosstalk in Diarrhea-Irritable Bowel Syndrome. Biomed Res Int. 2020; 2020: 3828249.
- Friedman ES, Li Y, Shen TD, Jiang J, Chau L, et al. FXR-Dependent Modulation of the Human Small Intestinal Microbiome by the Bile Acid Derivative Obeticholic Acid. Gastroenterology. 2018; 155: 1741-1752.e1745.
- Kunst RF, De Waart DR, Wolters F, Duijst S, Vogels EW, et al. Systemic ASBT inactivation protects against liver damage in obstructive cholestasis in mice. JHEP Rep. 2022; 4: 100573.
- Park JH, Iwamoto M, Yun JH, Uchikubo-Kamo T, Son D, et al. Structural insights into the HBV receptor and bile acid transporter NTCP. Nature. 2022; 606: 1027-1031.
- He S, Lin F, Hu X, Pan P. Gut Microbiome-Based Therapeutics in Critically Ill Adult Patients-A Narrative Review. Nutrients. 2023; 15.
- Song JH, Kim YS. Recurrent Clostridium difficile Infection: Risk Factors, Treatment, and Prevention. Gut Liver. 2019; 13: 16-24.
- Kelly CR, Allegretti JR. Review Article: Gastroenterology and Clostridium difficile Infection: Past, Present, and Future. Clin Infect Dis. 2023; 77: S463-s470.
- Weingarden AR, Chen C, Bobr A, Yao D, Lu Y, et al. Microbiota transplantation restores normal fecal bile acid composition in recurrent Clostridium difficile infection. American journal of physiology. Gastrointestinal and liver physiology. 2014; 306: G310-319.
- Bloom PP, Tapper EB, Young VB, Lok AS. Microbiome therapeutics for hepatic encephalopathy. J Hepatol. 2021; 75: 1452-1464.
- Bajaj JS, Salzman NH, Acharya C, Sterling RK, White MB, et al. Fecal Microbial Transplant Capsules Are Safe in Hepatic Encephalopathy: A Phase 1, Randomized, Placebo-Controlled Trial. Hepatology. 2019; 70: 1690-1703.
- Bajaj JS, Kassam Z, Fagan A, Gavis EA, Liu E, et al. Fecal microbiota transplant from a rational stool donor improves hepatic encephalopathy: A randomized clinical trial. Hepatology (Baltimore, Md.). 2017; 66: 1727-1738.
- Philips CA, Ahamed R, Rajesh S, Singh S, Tharakan A, et al. Clinical outcomes and gut microbiota analysis of severe alcoholassociated hepatitis patients undergoing healthy donor fecal transplant or pentoxifylline therapy: Single-center experience from Kerala. Gastroenterol Rep (Oxf). 2022; 10: goac074.
- Sharma A, Roy A, Premkumar M, Verma N, Duseja A, et al. Fecal microbiota transplantation in alcohol-associated acute-onchronic liver failure: an open-label clinical trial. Hepatol Int. 2022; 16:433-446.
- Horvatits T, Drolz A, Roedl K, Rutter K, Ferlitsch A, et al. Serum bile acids as marker for acute decompensation and acute-onchronic liver failure in patients with non-cholestatic cirrhosis. Liver Int. 2017; 37: 224-231.