
Journal of Gastroenterology Research and Practice
Research Article - Open Access, Volume 4
Curcumin nanoemulsions on the expression of fatty acid transporter proteins during hepatic steatosis
Beatriz Agame-Lagunes1; Alaina A Esperón-Rojas1; Peter Grube-Pagola2; Alfonso Alexander-Aguilera3; Rebeca Garcia-Varela4*; Hugo S García1*
1UNIDA, Tecnológico Nacional de México/Instituto Tecnológico de Veracruz. Miguel Ángel de Quevedo 2779, Veracruz, Ver. 91897, México.
2Universidad Veracruzana, Instituto de Investigaciones Medico Biológicas, Iturbide S/N, Veracruz, Ver. 91700, México.
3Universidad Veracruzana, Facultad de Bioanálisis. Iturbide S/N, Col. Centro, Veracruz, Ver. 91700, México.
4Carbone Cancer Center, University of Wisconsin School of Medicine and Public Health, Madison, Wisconsin, USA.
*Corresponding Author : Garcia-Varela R
Carbone Cancer Center, University of Wisconsin School of
Medicine and Public Health, Madison, Wisconsin, USA.
Email: garciavarela@wisc.edu
García HS
UNIDA, National Technological Institute of Mexico/Technological Institute of Veracruz, Miguel Ángel of Quevedo
2779, Veracruz, Ver. 91897, México.
Tel: +52 2291612351 & +52 2291614561;
Email: hugo.gg@veracruz.tecnm.mx
Received : Jan 08, 2024
Accepted : Feb 12, 2024
Published : Feb 19, 2024
Archived : www.jjgastro.com
Copyright : © Varela RG & Garcia B (2024).
Abstract
Introduction: Fructose intake of over 70 g/day is known to cause dyslipidemias, involving multiple metabolic pathways that can promote an excess of hepatic lipid synthesis as metabolic dysfunction-associated fatty liver disease (MAFLD), formerly known as non-alcoholic fatty liver disease. There is no current treatment that alleviates all the symptoms involved in this pathology.
Methods: O/W nanoemulsions, that were stable for 5 weeks, were prepared using a mono- and diacylglyceride emulsifier prepared by solvent-free glycerolysis; their effect was evaluated on a Wistar rat model with 30% fructose intake for 20 weeks and 5 weeks of treatment. qPCR was performed to determine the level of expression of fatty acid transporter proteins (FAPT 2, 5 and FABP).
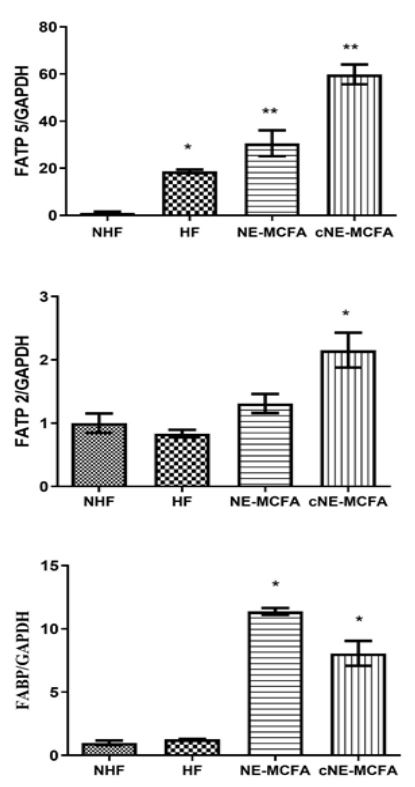
Results: Body weights and adipose tissue decreased significantly in experimental groups; liver tissue was comparable to the non-fructose (healthy control) group. Histological analysis showed a decrease in the presence of steatosis. Serological analysis reported a decrease in glucose and triglycerides in the NE-MCFA group (emulsifier without the active compound); however, both NE-MCFA and cNE-MCFA (emulsifier with curcumin) groups had decreased cholesterol, LDL-C, insulin, and the insulin resistance index (HOMA-IR), improved atherogenic values and transaminases ratio. The fatty acid transport protein, FATP 5, gene expression levels decreased in the NE-MCFA group, the FATP 2 increased in the cNE-MCFA group while the fatty acid binding protein, FABP, decreased.
Conclusion: These findings indicate the benefits that the use of nanoemulsions, stabilized with non-commercial emulsifiers, may have on MAFLD.
Keywords: Curcumin; Hepatic steatosis; Monoacylglycerides; Diacylglycerides; Medium chain fatty acids; Fatty acid transporter.
Abbreviations: MAFLD: Metabolic Dysfunction-Associated Fatty Liver Disease; O/W: Oil/Water; FATP: Fatty Acid Transporter Proteins; FABP: Fatty-Acid-Binding Proteins; NE: Nanoemulsion; HOMA-IR: Homeostatic Model Assessment; MAG: Monoacylglycerides; DAG: Diacylglycerides; MCT: Medium Chain Triglycerides.
Citation: Agame-Lagunes B, Esperón-Rojas AA, Grube-Pagola P, Alexander-Aguilera A, García HS, et al. Curcumin nanoemulsions on the expression of fatty acid transporter proteins during hepatic steatosis. J Gastroenterol Res Pract. 2024; 4(2): 1186.
Introduction
Ever since fructose was first manufactured industrially, its demand as an ingredient has increased; this has been driven by its lower cost and high sweetening power; causing the current average consumption of fructose to exceed 70 g/day. Recently published research reported a strong relationship between the excessive consumption of this monosaccharide and metabolic disorders [1]. The absorption of fructose, between 5-50 g, in the enterocyte villi is mediated by the GLUT-5 transporter, which crosses from the enterocyte to the portal vein by means of the GLUT-2 transporter. Although its absorption is slower compared to that of glucose, its metabolism occurs rapidly in the liver and is transported to the hepatocyte by the GLUT-2, 5, 8 or 9 transporters, decreasing the stimulation for insulin release [2].
Fructose metabolism is facilitated by fructokinase to produce fructose 1-P, which is metabolized by an aldolase to Dihydroxyacetone Phosphate (DHAP) and glyceraldehyde. DHAP can enter glycolysis directly, but glyceraldehyde must be phosphorylated by triose kinase. When there is an increased intake of fructose, both trioses are converted to acetyl CoA and thus synthesize fatty acids, leading to an accumulation and use of the trioses for the formation of glycerol and fatty acids [3]. Fructose intake is highly related to Metabolic-Associated Fatty Liver Disease (MAFLD), previously known as Non-Alcoholic Fatty Liver Disease (NAFLD), which is a set of metabolic imbalances that may also include genetic and epigenetic factors, starting with hepatic steatosis [4], which involves the excessive absorption of triacylglycerides into the hepatocyte. Once the intestinal capacity is exceeded, fructose is transported by the portal vein to the liver where 80-70% is metabolized. Therefore, a high fructose diet induces de novo synthesis and accumulation of triacylglycerides and decreases in the β-oxidation of free fatty acids [5,6]. This pathology affects over 20% of the population [7]. MAFLD is considered the hepatic component associated with the metabolic syndrome because 80% of the obese population and diabetes mellitus 2 patients have it [2]; the association of MAFLD with other pathologies has increased in prevalence, mortality and morbidity [8]. The 2018 National Health and Nutrition Survey reported over 70% obesity in the adult population and 35% in infant population [1]. Also, the prevalence of ≈40% MAFLD amongst Latino populations has been estimated to be the highest [9]. Since one of the main causes for the development of fatty liver is an unhealthy lifestyle, a complete modification of eating habits and prevention of sedentary lifestyles are suggested; however, the lack of discipline in the population undermines the benefits of following these recommendations. For this reason, the use of complementary therapies based on encapsulated functional compounds that improve their bioavailability has been suggested [10,11]. Curcumin, derived from the rhizome of Curcuma longa (diferuloylmethane), is recognized for its beneficial properties as an antioxidant [12], anti-inflammatory [13], antiparasitic [14], antifibrotic, anticancer, and is considered a good hepatoprotective agent [15]. Nevertheless, this compound has the disadvantages of being poorly permeable in the gastrointestinal tract, chemically unstable and its effects are dose-related because of its lipophilic nature. Therefore, the use of carrier systems has been implemented to avoid using high doses of curcumin, although it holds the status of Generally Recognized as Safe (GRAS) at doses up to 2 g curcumin encapsulated/kg body weight [16,17]. Nanoemulsions (NE) are structures mainly composed of oil, water, emulsifier(s) and the compound of interest to be encapsulated. One of the benefits of this system is the protection that NE structures provide to the compound from degradation by avoiding contact with oxygen or hydroxyl ions. It is important to mention that the smaller the particle size, the higher the degradation rate of the compound [18]. The efficiency of NE to be absorbed by enterocytes is depend on their diameter (20-200 nm); also, the chain length of the oil incorporated is highly related to the bioaccessibility of the bioactive [19]. The advantage of using a designed emulsifier may represent greater abilities and controlled qualities that can be conferred to the system. The use of Monoacylglycerides (MAG) and Diacylglycerides (DAG) has the additional benefit of not requiring hydrolysis of fatty acids by pancreatic lipases, providing greater absorption by enterocytes for subsequent passage across the plasma membrane. This can be attained by either passive diffusion or by fatty acid transporters [20]. Medium Chain Fatty Acids (MCFA) such as caproic, caprylic, and capric acids contribute to the formation of ketone bodies. When consumed in the diet, they are rapidly absorbed by the portal system and undergo β-oxidation in the liver; in turn, this increases acetyl-CoA concentrations [21]; may function as inflammatory mediators, antioxidants, increase the translation of genes related to fatty acid metabolism, and are involved in the decrease of the mTORC1 pathway [22,23]. In others such as caproic acid activate 5’AMP-Activated Protein Kinase (AMPK) that inhibits glycogen synthesis and lipolysis of adipose tissue, their concentration decreases in some pathologies [24]. The Fatty Acid Binding Protein (FABP1) is involved in fatty acid homeostasis. Although it has several isoforms, they all share similar structures: a β domain and a section for lipophilic ligands, i.e., they function as chaperones to mediate lipid transport to organelles within the cell. They are located in the cytoplasm of hepatocytes and the outer mitochondrial membrane; each molecule binds to two long-chain fatty acid molecules independently and form favorable hydrophobic bonds. This protein interacts with ligands, binding with acyl CoA, bile acids, fatty acid metabolites and possibly MAG [25]. The fatty acid transport protein, FATP, or solute carriers 27A, comprises a family of 6 isoforms, being isoforms 2 and 5 those expressed in the liver [26]. Based on previous research from our group, where favorable results were reported, the aim of this research was to evaluate possible beneficial effects of these newly designed NE as hepatoprotective components, after 20 weeks of induction of hepatic steatosis. The diets contained high fructose intake in rats. In this work, a longer period of fructose intake was applied and expression levels of some fatty acid transporter proteins were analyzed as an attempt to clarify some questions that were raised in our previous report [27].
Materials and methods
Reagents
High purity curcumin (≥98%) was purchased from LKT Laboratories (St. Paul, MN, USA). Medium chain triglycerides were obtained from Swanson Health Products (Fargo, ND). A Zetasizer Nano-ZS90 equipment with dynamic light scattering (Malvern Instruments Inc., Worcestershire, UK) was used. Golden Bell brand glycerol (Mexico City, Mexico), Kolliphor® EL, caproic, caprylic and capric fatty acids with purity ≥98%, were all obtained from Sigma-Aldrich (Mexico City, Mexico); Novozym 435 (lipase from Candida antarctica fraction B) was a gift from NOVO (Barcelona, Spain). KRYSTAR® 300 fructose (≥99% purity) was purchased from TATE & LYLE (London, UK).
O/W curcumin Nanoemulsions (NE)
The emulsifier was prepared and purified according to our previous publication [28]; briefly, using a combination of caproic (C6:0), caprylic (C8:0) and capric (C10:0) acid in a 1:1:1 ratio w/w/w, 5% Novozym 435 lipase and 10% of 3Å molecular sieves; the w% of lipase and molecular sieves was set in accordance to total substrates. The reaction mix was placed in a Thermo MaxQ 4450 orbital shaker set at 300 rpm and 50°C for 30 minutes. The O/W curcumin NE were formulated according to the procedure described in our previous publication [27]; briefly, the oil and aqueous phases were prepared separately with their components described in Table 1. Both phases were placed in a Barnstead/Labline Aquawave 9376 (Singen, Germany) ultrasonic bath for 30 minutes, then mixed and positioned in an Ultra-Turrax T25 homogenizer for 3 minutes set at 20,000 rpm with 1-minute intervals. Finally, to obtain the NE a Branson Digital S-450D ultrasonic device was used at 20% amplitude for 3 minutes with 1-minute intervals in an ice bath. Polydispersity index (PDI) and the stability of the average particle size (Zaverage) were measured with a Zetasizer Nano ZS90 (Malvern Instruments Inc., Worcestershire, UK) with dynamic light dispersion at 90° scattering angle at 25°C.
Fructose-induced hepatic steatosis
Based on our previous research [27], the experimental period was extended to 20 weeks for the induction of hepatic steatosis with 30% fructose, and 5 weeks of further treatment. Briefly, 20 21-week-old male Wistar rats, supplied by the Cristóbal Colón University bioterium, were kept in individual stainless-steel cages to provide adequate housing, with controlled temperature in 12 h/12 h light/dark cycles. Maintenance and handling conditions were performed in accordance with the guidelines provided by the Guide for the Care and Use of Laboratory Animals of the National Research Council, eighth edition (2011). Animals were allowed to adapt for one week with a standard diet (Laboratory Rodent Diet 5001, Lab Diet) and water ad libitum.
After the adaptation period, the animals were distributed in two groups: a control group without fructose (n=5) and a second control group with fructose (n=15). Both groups consumed the standard diet; however, for 20 weeks, the control group with fructose (HF) was provided with water with 30% fructose in their container, in contrast to the control group which only had water without fructose (NHF). After 20 weeks of induction, the HF group was divided into three groups: a) Sick control group (HF) (n=5); b) curcumin NE with MAG and DAG enriched with MCFA (cNE-MCFA) (n=5); and NE with MAG and DAG enriched with MCFA without curcumin (NE-MCFA) (n=5). Treatments corresponding to each group were administered by gastric cannula (20 mg curcumin or NE/kg body weight of the rat) and the HF and NHF groups were provided with water/glycerol (50/50) for 5 weeks.
Nutritional status measurements
In order to determine and compare the nutritional status of the animals in each group, the initial weight of each of the study subjects was recorded; this parameter continued to be monitored for 25 weeks. In the same manner, the consumption of food and liquid, with and without 30% fructose, was also monitored; therefore, the caloric equivalents of each intake were also estimated.
Serological parameters
Blood samples were obtained at two different stages by puncture of the rats’ lateral vein: the first stage was after 20 weeks of induction with 30% fructose, and the second stage was at the end of the experimental phase at 25 weeks. In both cases, a period of 8-10 h fasting was maintained. Blood samples were centrifuged at 1086xg for 10 minutes to obtain the serum; samples with hemolysis were discarded. Glucose, cholesterol, triacylglycerides, Aspartate Aminotransferase (AST), Alanine aminotransferase (ALT), LDL and HDL content were determined by spectrophotometric techniques; serum insulin quantification was performed by an immunoenzymatic assay (MEXLAB GROUP 6001011). To calculate the insulin resistance index (HOMA-IR), the following equation was used:
Hepatic histology
To determine liver tissue damage caused by the intake of 30% fructose, tissue samples were stored in 10% formaldehyde and embedded in paraffin for hematoxylin and eosin staining (H&E). Images were obtained using an Olympus BX51 microscope (Tokyo, Japan) equipped with a digital camera. A semiquantitative analysis of lipid droplets was performed in 20 fields and observed at 10X and 20X, considering a positive field to have at least 50% of lipid vacuoles present.
Gene expression
Liver tissue was pulverized by adding liquid nitrogen in a stainless-steel mortar, specially adapted to homogenize biological samples, and stored for later use at -20°C; RNA was extracted according to the instructions of the Trizol reagent from Sigma-Aldrich; purity was determined in a SmartSpec™3000 equipment.
A commercial QuantiNova™SYBR Green RT-PCR kit from Qiagen N.V. (Hilden, Germany) was used to perform qPCR according to the manufacturer’s guidelines; the QuantStudio™ RealTime PCR software v 1.3 was employed, which uses the 2-ΔΔCT method. The primers used are listed in Table 2.
Data analysis
Statistical analysis was performed using Minitab® v. 18 software (Minitab Inc., State College, PA); ANOVA and paired comparison using Tukey’s test (p<0.05 or p<0.01) was made with a confidence level greater than 95% to be considered statistically significant. Results were expressed as mean ± standard deviation.
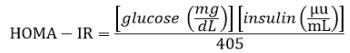
Results
Physical characteristics of curcumin O/W NEs with MAG and DAG enriched with MCFA as emulsifier
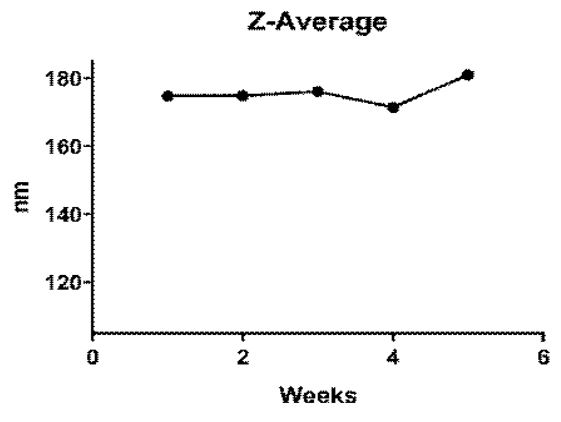
NE with mean particle sizes of 174.8±1.11 nm and a polydispersity index (PDI) of 0.195±0.076 were obtained and stored at 4°C for 5 weeks; these NE showed stability during this time with a variability in particle size of 175.66±3.44 nm (Figure 1) and a PDI of 0.0822±0.026.
Table 1: Composition of O/W curcumin NE with MAG and DAG enriched with MCFA.
| Oil phase | |
|---|---|
| Component | Concentration/ratio |
| Curcumin | 2.5 mg/g NE |
| Medium chain oil | 3% |
| Kolliphor® EL | 1% |
| Aqueous phase | |
| Component | Concentration/ratio |
| Milli-Q water | 49% |
| Glycerol | 50% |
| MAG and DAG enriched with MCFA | 1% |
Table 2: Primers designed for gene expression monitored by means of RT-PCR.
| Gene | Forward (5´-3´) | Reverse (5´-3´) |
|---|---|---|
| FATP-5 | TTCAGGGACCACTGGACTTC-CAAA | ACCACATCATCAGCTGTTCTCCCA |
| FATP-2 | AGTACATCGGTGAACTGCTTC-GGT | TGCCTTCAGTGGAAGCG-TAGAACT |
| FABP | TCTGCTTCACCGGCCTAAA | AGATTCGACCATCCTTTGTCATG |
| GAPDH | ATGTGTCCGTCGTGGATCTGA | TTGAAGTCGCAGGAGACAACCT |
Table 3: Weight, water, and feed consumption; caloric equiva- lent monitored in Wistar rats after 20 weeks of induction with 30% fructose.
| NHF | HF | |
|---|---|---|
| Weight | ||
| Basal (g) | 203.6±22.0 | 198.3±36.5 |
| 10 weeks (g) | 337±7.8 | 318±13 |
| 20 weeks (g) | 398±17 | 372±23 |
| Consumption of liquid, food and its caloric equivalents | ||
| Liquid ML/day | 110±15 | 110±11 |
| Liquid ML/day/100 g bw | 32±6.9 | 36±8.1 |
| Equivalent calories | 0±0 | 43.60±9.76* |
| g/Day/100 g bw | 17±3.7 | 9.2±2.9* |
| Equivalent calories | 60±13 | 31±9.9* |
Data Means ± SD of rats in the control group without fructose NHF (n=5) and control group with 30% fructose HF (n=15); *P<0.05.
Induction of hepatic steatosis by correlating nutritional condition and serologic variables
From the beginning of the experimental phase, no significant difference between the subjects in the NHF and HF groups, with respect to weight, was found. This trend continued at weeks 10 and 20, despite the HF group having a 30% fructose intake in water (Table 3); the NHF group had a 65% increase in weight at week 10, while the HF group had a 60% weight gain in the same period. By week 20 the NHF group had a 95% weight gain compared to their initial weight and the HF group had an 88% increase compared to their initial weight. Although weight increase was observed when each group was analyzed, there were no significant differences found between them. Furthermore, data on feed, water, water with 30% fructose and their corresponding caloric equivalents were also monitored. Although there was no statistical difference in liquid intake between the groups, there was a difference between the caloric equivalents, with the HF group displaying a significant increase. Feed intake was higher in the NHF group as well as the corresponding caloric equivalent.
To verify that the study subjects were in a hepatic steatosis state, caused by the 30% fructose intake, a sampling to measure different biochemical parameters was made by blood collection from the tail lateral vein. Rats were previously fasted for 8-10 hours. As it can be seen in Table 4, there was no significant difference in glucose levels between the NHF and HF groups; however, in the HF group there was a 60% increase in cholesterol values and triglycerides above 50%, a three-fold increase in LDL and there was no significant difference in the insulin values; the AST/ALT ratio was less than 1, indicating that this group was indeed in steatosis.
Evaluating the effect of treatments on hepatic steatosis
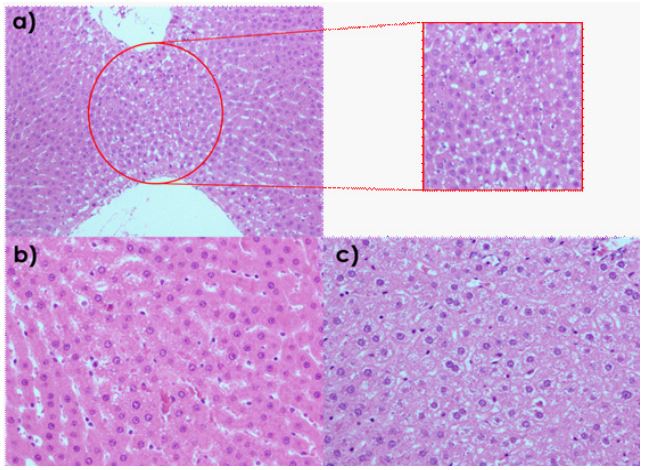
Once it was confirmed that the HF group was in hepatic steatosis, this group was subdivided for administration of the NE with and without curcumin in the corresponding groups (NEMCFA and cNE-MCFA), and the sick control group. Each treatment was administered orally through a gastric cannula. The effects of the treatments on weight gain were 89% in the cNE-MCFA group, 97% in the NE-MCFA group and the TE group had an increase of more than 111% (Table 5). Regarding liver weight, the HF group displayed a decrease in liver weight compared to the NHF, NE-MCFA and cNE-MCFA groups. Adipose tissue was also collected for weight comparison between each group, with a 39% decrease in the NE-MCFA group and a 51% decrease in the cNE-MCFA group when compared with the TE group. Although the hepatosomatic index was higher in the groups treated with NE with and without curcumin, the extent of steatosis determined after the histological analysis in liver tissue was lower in both cases (Table 5 and Figure 2). A semiquantitative visual analysis of lipid droplets was performed in 20 completely randomized fields; observations were carried out at 10X and 20X. A field was considered positive when 50% of lipid droplets were found. The NHF group had a 1% development of steatosis, while the group with 30% fructose without treatment developed over 7% of steatosis. The NE-MCFA and the cNE-MCFA groups had lower extents compared to the HF group of 4% and 0% respectively; however, the cNE- MCFA group showed 40% of glycogen granules.
To compare in serum the effects of NE-MCFA and cNE-MCFA treatments on fructose-induced hepatic steatosis, after 5 weeks of treatment, rodents were fasted for 8-10 h prior to euthanasia and blood was collected by cardiac puncture. When comparing the levels at week 25 (Table 6) with the values at the week before treatment started (Table 4), glucose values decreased only in the NE-MCFA group over 17%. Cholesterol levels decreased significantly in both NE treatments by over 38% (NE-MCFA) and 29% (cNE-MCFA); these values were comparable to those from the HF group. Triacylglycerides content decreased only in the NE-MCFA group. The AST/ALT ratio was less than 1 only in the HF group. LDL of the cNE-MCFA group decreased to values similar to the NHF group (64%) and the NE-MCFA group declined by 49%. Insulin and HOMA-IR levels were lower in the groups that received the NE treatments with and without curcumin.
As depicted in Figure 3, there was an overexpression of all fatty acid transporters (FATP 5, FATP2 and FABP), however, the NE-MCFA and cNE-MCFA groups were further increased compared to NHF and HF.
Table 4: Serological parameters in the NHF control group with- out fructose and the HF control group with 30% fructose, after 20 weeks.
| Parameter | NHF | HF |
|---|---|---|
| Glucose (mg/dL) | 137 ±4.7 | 144±13 |
| Cholesterol (mg/dL) | 47±9.5 | 76±5.7* |
| Triacylglycerides (mg/dL) | 72±4.7 | 110±19* |
| Aspartate aminotransferase (AST) (U/L) | 76±1.5 | 23±2.5* |
| Alanine aminotransferase (ALT) (U/L) | 31±3.5 | 91±11* |
| LDL (mg/dL) | 57±6 | 170±8.5* |
| Insulin (μUI/mL) | 4.8±0.11 | 3.9±0.66 |
| HOMA-IR | 1.6±0.055 | 1.44±0.25 |
| Data means ± SD of rats in the NHF (n=5) and the HF (n=15); *P<0.05. | ||
Table 5: Effect after 5 weeks of treatment for the induction of hepatic steatosis in rats with 30% fructose for 20 weeks.
| Parameter | NHF | HF | NE-MCFA | cNE-MCFA |
|---|---|---|---|---|
| Final weight (g) | 419.33±8.7 | 410±9.0 | 380±14* | 387±9.1* |
| Weight gain (g) | 208±9.4 | 212±0.8 | 184±1.5* | 189±19 |
| Liver weight (g) | 12.67±0.6 | 9.80±1.8* | 11.80±1.3 | 13.80±0.8 |
| Adipose tissue weight (g) |
20.33±1.5 | 25.67±2.1* | 15.67±2.1** | 12.67±0.6** |
| Hepatosomatic index | 3.02±0.1 | 2.19±0.5 * | 3.21±0.3 | 3.62±0.2 |
| Steatosis | 1% | 7% | 4% | 0% |
| Glycogenization | 0% | 0% | 0% | 40% |
|
Data means ± SD of rats in the control group without
fructose NHF (n=5), control group with 30% fructose HF
(n=5), NE with MAG and DAG enriched with MCFA without the curcumin active compound NE-MCFAs (n=5) and curcumin NE with MAG and DAG enriched with MCFA cNE- MCFAs (n=5); *P<0.05, **P<0.0001. |
||||
Table 6: Serological parameters in each of the NHF, HF, NE-MC- FA and cNE-MCFA groups after the corresponding treatments, for 25 weeks.
| Parameter | NHF | HF | NE-MCFA | cNE-MCFA |
|---|---|---|---|---|
| Glucose | 143±6.5 | 178±3.1** | 111±10* | 180±6.5** |
| Cholesterol | 57±4.5 | 78±12* | 48±5.3 | 55±4.5 |
| Triacylglycerides | 91±12 | 150±12** | 100±9.6 | 143±16** |
| AST/ALT | >1 | <1* | >1 | >1 |
| LDL | 54±3.5 | 204±10** | 86±11* | 61±2.0 |
| HDL | 41±3.6 | 34±4.7 | 52±9.2* | 47±3.5* |
| Insulin | 4.8±0.3 | 5.1±0.7 | 3.8±0.1* | 3.4±0.3** |
| HOMA-IR | 1.7±0.1 | 2.3±0.3* | 1.1±0.1** | 1.5±0.2 |
|
Data means ± SD of rats in the control group without
fructose NHF (n=5), control group with 30% fructose HF (n=5), NE with MAG and DAG enriched with MCFA without the active compound NE-MCFAs (n=5) and curcumin NE with MAG and DAG enriched with MCFA cNE-MCFAs (n=5); *P<0.05, **P<0.0001 |
||||
Discussion
The present study was intended as a second phase of our previous research regarding the study of the effect of curcumin NE with a non-commercial emulsifier that enhanced its bioavailability [27]. The designed emulsifier consisted of MAG and DAG enriched with MCFA. Our previous published research was performed using the same model for only 10 weeks of induction of hepatic steatosis, with 30% fructose intake and further two weeks with NE treatment. Beneficial results in several serological parameters were obtained, except in triacylglycerides values that were elevated in groups that were treated with curcumin; therefore, this was one of the main variables considered to determine if by increasing treatment time, harmful effects of hepatic steatosis could be reduced.
Several experiments with fructose feeding models indicate that high daily fructose consumption causes hyperlipidemia, Insulin Resistance (IR) and non-alcoholic fatty liver disease [29,30]. Nevertheless, these results are ambiguous most likely attributed to variable environmental and genetic factors. A comparison between non-obese patients with fatty liver disease and obese patients without fatty liver disease has been reported, showing similar levels of dyslipidemia and hypertriglyceridemia but with higher hemoglobin levels in patients with fatty liver. Another effect fructose exerts on the liver is energy depletion that causes its catabolism inducing adenylate kinase to replenish ATP and AMP from 2 ADP. AMP is then degraded to uric acid, decreasing NO with a concomitant reduction in muscle tone, activating mitochondrial oxidative stress, which induces fatty acid synthesis [31].
It has been suggested that healthier lifestyles that include a balanced diet and exercise, combined with the use of drugs such as metformin, can contribute to the reduction of hepatic steatosis [32]; however, adverse effects have been reported in the chronic use of drugs [33]. In the same way, the general population is often unable to sustain a healthy lifestyle. For this reason, the application of alternative/complementary therapies, using bioactive compounds like curcumin, with antioxidant and hepatoprotective benefits, has been proposed. However, curcumin has low water solubility caused by its lipophilic nature; in consequence, delivery systems as O/W nanoemulsions have been studied to increase its bioavailability [34].
Here, NE particle sizes were smaller than 200 nm, and the PDI was below 1, suggesting that stability of the system was conferred by the formulated emulsifier, with a variability under 10 nm in 5 weeks of storage at 4°C, as reported in our previous research [27].
Regarding the Wistar rat model, provided with 30% fructose in their drinking water for 20 weeks to induce hepatic steatosis, despite a constant increase in animal weight compared to their baseline, there was no significant difference between the fructose and non-fructose groups. This is comparable to values reported by Miranda et al. [35], where an increase in body weight was expected; yet it was observed that when using fructose there was no evident change. This parameter is still controversial since some authors report a difference in weight between groups when fructose was used [31,36]; however, others did not share this finding; such is the case of this research. Nevertheless, after receiving the corresponding treatments, there was a decrease in weight in the NE-MCFA and cNE-MCFA groups; these results are comparable to our previously published report supporting the anti-obesogenic effect that MCFA and curcumin can provide in a dose-dependent manner [27]. Similarly, Maithilikarpagaselvi et al. [37] using 40 male Wistar rats, provided with 60% fructose in the feed for 10 weeks, together with a treatment of 200 mg/kg body weight of curcumin, reported a decrease in body weight. Additionally, Lee et al. [38] described a reduction in body weight after providing curcumin NE (1.64±0.01 µg/mg) to BALB/c mice.
Although there was no significant difference in the consumption of liquid with and without fructose during 20 weeks, the caloric equivalent increased in the HF group. This was attributed to the added caloric contribution when consuming the fructose in the rats’ drinking water, as opposed to what was obtained in the consumption of food. The NHF group ingested more feed in g/day-100 g body weight of the rat, obtaining higher values of caloric equivalent in relation to what was established in the nutritional information of the food provided. The decrease in food consumption in the HF group is attributed to the fact that rats first satisfy their liquid requirements and then their solid food needs; however, by covering this caloric need with the consumption of fructose in the liquid, they no longer require as much food consumption; therefore, there was a compensatory calorie effect [39]. These results are comparable to those reported by Spruss et al [32] who provided C57BL/6J mice with a 30% fructose solution for 8 weeks and showed an increase in caloric equivalent but no difference in body weight.
To determine the serological values, samples were analyzed using commercial kits, following the supplier’s instructions. Glucose values after 20 weeks of induction showed no difference between the groups with 30% fructose (HF) and without fructose (NHF). However, after providing the corresponding treatment, this variable decreased in the NE-MCFA group. These results align with data reported by Watanabe et al. [40]. where they mention that MCFA besides decreasing body weight, also, when ingested before food, produced lower postprandial glucose levels. This may be caused by alternative energy sources being used, such as ketone bodies produced by ingestion of this type of fatty acid, and that lipid consumption is not affected. As for cholesterol values, there was a significant difference between the groups with and without fructose after 20 weeks of induction, as it was reported by Oseini et al. [41]. Conversely, after treatment (week 25) these values decreased in the NE-MCT and cNE-MCFA groups to levels comparable with the healthy control group (NHF). This may be attributed to cholesterol regulation, since there are 3 metabolic pathways to maintain a state of homeostasis: via biosynthetic, exogenous and bile release pathways. It has been reported that MCFA increase the expression of a cluster of proteins that transport dietary sterols to prevent their accumulation in the liver and small intestine; therefore, MCFA promote cholesterol efflux and increase the expression of sterol transport proteins, thus indirectly decreasing serum cholesterol concentrations [42]. In the HF groups, triacylglycerols content increased after 20 weeks of induction; but, after receiving the corresponding NE treatments, there was only a decrease in the NE-MCFA group similar to what we previously reported [27]. In a study conducted by Chien et al. [43], a systematic review and meta-analysis of randomized controlled trials was conducted to investigate the effect of curcumin in patients with polycystic ovary disease on lipid profile; 500 mg of curcumin were administered from one to three times a day, during a period of 6 to 12 weeks. The authors determined that curcumin improved HDL and total cholesterol levels but not LDL or triacylglycerols content; also, in a randomized controlled trial in older adults with obesity, the effects of fish oil on cardiovascular parameters were determined. No additional beneficial results were observed when a supplementation of fish oil in combination with 160 mg/day of curcumin were employed [44]. This is consistent with our obtained data. Although, curcumin has numerous reports indicating a lowering of triacylglycerols in serum, as published by Thota et al. [45]. In their research a double-blind, placebo-controlled study was conducted where the effect of curcumin (2 capsules of 500 mg) or with omega-3 polyunsaturated fatty acids (2 capsules of fish oil equivalent to 1.2 g DHA and EPA) on blood lipids for 12 weeks were assessed. A preservation of glucose levels was observed, improved insulin sensitivity in the curcumin group and, to a lesser extent, in the combined curcumin+fish oil group. Similarly, the curcumin and curcumin + fish oil group improved triacylglycerols levels. Nonetheless, the group that showed a greater improvement in serum parameters was the group that consumed fish oil only. This also suggests the importance of the dosage and the oil involved at the time of administration in the study subjects. As can be seen in our data, there was an improvement in the AST/ ALT ratio after the treatments which suggests hepatic restoration. The effect of 14-day consumption of curcumin nanoparticles (50 mg/kg body weight/day) on hepatoxicity caused by cisplatin in rats, has been reported, indicating an improvement in transaminases and in the histological structure of the hepatocyte [46]. Transaminases are normally found in the cytoplasm of the hepatocyte; when liver damage occurs, a leaking occurs due to a loss of membrane integrity. Hence, curcumin can reorganize and heal hepatic damage, since it is capable of reducing liver transaminase activity by enhancing liver tissue structure [46]. There was an amelioration in LDL and HDL lipoprotein levels, this indicates an improvement in the atherogenic index of both NE-MCFA and cNE- MCFA groups due to the cardioprotective effects of curcumin given its lipid-lowering properties, although it may not have counteracted high triglyceride levels, but it could have performed its function by activating PPAR-γ and thus suppress the LDL receptor gene and decrease its concentration [47].
Insulin resistance is considered as a predictor for the progression to type-2 diabetes mellitus, since an insulin deficit produces a deficiency in glucose homeostasis. According to previously published research, this improvement in insulin resistance may be attributed to a decrease in NFκB activity, an increase in antioxidant transcription factor and activation of AMPK, sites in which curcumin functions as a mediator; also, MCFA promote lipid oxidation and increase insulin sensitivity; for this reason, its use is recommended for weight loss [48]. Thota et al. [45] reported that there was an improvement in insulin sensitivity in groups that were fed with curcumin, fish oil and the combination of curcumin with fish oil.
In order to perform the histological analysis, livers were first weighed and data recorded. Weights were comparable in the NE-MCFA and cNE-MCFA treatments and the NHF group. In the same manner, adipose tissue weight was recorded. Body fat of the cNE-MCFA and NE-MCFA groups decreased by approximately 51% and 39%, respectively, compared to the HF group, with 30% fructose intake without treatment; however, when comparing the ratio between body weight and liver size, the cNE-MCFA group was found to be higher. According to the histological analysis there was a decrease in lipid vacuoles in the NE-MCFA group; nevertheless, in the cNE-MCFA group glycogenization process, an approximate 51% decrease in adipose tissue was recorded. Shen et al. [49] used Sprague Dawley rats subjected to chronic stress conditions for 12 weeks. The model was provided with curcumin at high, medium, and low doses (60, 30 and 15 mg/kg curcumin, respectively). Several effects were found, i.e., at high and medium doses serum glucose decreased and increased insulin. Meanwhile, at low doses glycogen values increased; these results may be attributed to the fact that insulin contributes to the decrease of non-esterified fatty acids, promoting glycogen storage and lipolysis inhibition. Also, Atia et al. [50] administered curcumin nanoparticles, of sizes smaller than 100 nm, to male Swiss albino mice, that were exposed to acrylamide. As a result, a decrease in lipid vacuoles was observed. It must be noted that in this research, despite having used curcumin in a nanocarrier system to improve its bioavailability, an overaccumulation of glycogen was attained in cells because, as reported by Hengist et al. [51], there may have been an overexpression of hepatic glycosidase. This could have resulted in increased production of glycogen storage sites. An elevation in triacylglycerols, together with the facility conferred by curcumin to elevate hepatic glycogen synthesis by inhibiting the glycogen synthase kinase 3b (GSK-3), favored higher triglyceride and glucose levels as it can be observed in our data [49]. Because FATP can function as an Acyl-Coa Synthase (ACS), this causes an increase in the uptake of fatty acids because it supports the creation of a concentration gradient. This means that increasing its level of expression will raise the uptake and activation of fatty acids, while FABP is bound to the non-esterified fatty acids for their subsequent oxidation or esterification. It has been observed that when esterification is hindered, accumulation of fatty acids in hepatocytes is produced [52]. The use of non-commercial emulsifiers allowed the formulation of nanoemulsions with sizes smaller than 200 nm with a stability longer than 4 weeks. The preparation allowed the decrease of the serological parameters associated to hepatic steatosis by promoting the influx of MCFA. In the same way, nanoencapsulated curcumin produced a hepatoprotective effect; however, this effect was lower in serum triacylglycerols levels, that could be attributed to an antagonistic effect between curcumin and the emulsifier which, as it contains fatty acids with different chain lengths, does not allow so much flexibility in the emulsion and thus limits the release of the active compound [53].
Conclusion
To conclude, the reported findings indicate the benefits of using NE as carriers of bioactive compound. Stable formulations were obtained using non-commercial emulsifiers, such as MAG and DAG. The NE containing curcumin produced an hepatoprotective effect; however, more research is required in order to fully understand the effect on additional parameters, for the potential use of this strategy as a co-adjuvant treatment for MAFLD.
Declarations
Ethics approval and consent to participate: Institutional Review Board Statement: This work was approved for animal handling by the institutional board of research on October, 26 2018 (CI-ITVER/10/2018).
Funding: The authors gratefully appreciate the Financial support of the Coveicydet of Veracruz through the Grant 14 2206.
Conflict of interest: The authors declare no conflict of interest.
Acknowledgements: The authors gratefully appreciate the financial support of the COVEICyDET of Veracruz through the grant 14 2206.
References
- Ponce-López ML, Zarco Villavicencio A, Robles-Aguirre BA, Cardoso-Gómez MA. Consumo de fructosa y sus implicaciones para la salud Fructose consumption and its implications for health. Rev Med e Investig. 2020; 8: 38-44.
- Loza-Medrano SS, Baiza-Gutman LA, Ibáñez-Hernández MÁ, Cruz-López M, Díaz-Flores M. Molecular alterations induced by fructose and its impact on metabolic diseases. Rev Med Inst Mex Seguro Soc. 2019; 56: 491-504.
- Gugliucci A, Rodríguez-Mortera R. Fructosa, un factor clave modificable en la patogenia del síndrome metabólico, la esteatosis hepática y la obesidad. Rev Medica del Uruguay. 2020; 36: 418-30.
- Gallego-Durán R, Albillos A, Ampuero J, Arechederra M, Bañares R, et al. Metabolic-associated fatty liver disease: From simple steatosis toward liver cirrhosis and potential complications. Proceedings of the Third Translational Hepatology Meeting, organized by the Spanish Association for the Study of the Liver (AEEH). Gastroenterol Hepatol. 2022; 45: 724-734.
- Muriel P, López‐Sánchez P, Ramos‐Tovar E. Fructose and the liver. Int J Mol Sci. 2021; 22: 6969.
- Ahadi M, Molooghi K, Masoudifar N, Namdar AB, Vossoughinia H, et al. A review of non-alcoholic fatty liver disease in nonobese and lean individuals. 2021; 36: 1497-1507.
- Arab JP, Dirchwolf M, Álvares-da-Silva MR, Barrera F, Benítez C, et al. Latin American Association for the study of the liver (ALEH) practice guidance for the diagnosis and treatment of non-alcoholic fatty liver disease. Ann Hepatol. 2020; 19: 674-690.
- Rojas YAO, Cuellar CLV, Barrón KMA, Arab JP, Miranda AL. Nonalcoholic fatty liver disease prevalence in Latin America: A systematic review and meta-analysis. Ann Hepatol. 2022; 27: 100706
- Bernal-Reyes R, Icaza-Chávez ME, Chi-Cervera LA, Remes-Troche JM, Amieva-Balmori M, et al. Prevalence and clinical-epidemiologic characteristics of a Mexican population with metabolic (dysfunction) associated fatty liver disease: An open population study. Rev Gastroenterol Mex. 2023; 88: 199-207.
- Stefan N, Häring HU, Cusi K. Non-alcoholic fatty liver disease: Causes, diagnosis, cardiometabolic consequences, and treatment strategies. Lancet Diabetes Endocrinol. 2019; 7: 313-324.
- Singh S, Osna NA, Kharbanda KK. Treatment options for alcoholic and non-alcoholic fatty liver disease: A review. World J Gastroenterol. 2017; 23: 6549-7650.
- Lin X, Bai D, Wei Z, Zhang Y, Huang Y, et al. Curcumin attenuates oxidative stress in RAW264.7 cells by increasing the activity of antioxidant enzymes and activating the Nrf2-Keap1 pathway. PLoS One. 2019; 14: 1-13.
- Peng Y, Dong B, Jiang Y, Yu L, Chen Z, et al. Anti-inflammatory effects of curcumin in the inflammatory diseases: Status, limitations and countermeasures. Drug Des Devel Ther. 2021; 15: 4503-25.
- Rai M, Ingle AP, Pandit R, Paralikar P, Anasane N, et al. Curcumin and curcumin-loaded nanoparticles: Ant pathogenic and antiparasitic activities. Expert Rev Anti Infect Ther. 2020; 18: 367-79.
- Farzaei MH, Zobeiri M, Parvizi F, El-Senduny FF, Marmouzi I, et al. Curcumin in liver diseases: A systematic review of the cellular mechanisms of oxidative stress and clinical perspective. Nutrients. 2018; 10: 855.
- Jantawong C, Priprem A, Intuyod K, Pairojkul C, Pinlaor P, et al. Curcumin-loaded nanocomplexes: Acute and chronic toxicity studies in mice and hamsters. Toxicol Reports. 2021; 8: 1346-1357.
- Ipar VS, Dsouza A, Devarajan PV. Enhancing Curcumin Oral Bioavailability Through Nanoformulations. Eur J Drug Metab Pharmacokinet. 2019; 44: 459-480.
- Kharat M, Aberg J, Dai T, McClements DJ. Comparison of Emulsion and Nanoemulsion Delivery Systems: The Chemical Stability of Curcumin Decreases as Oil Droplet Size Decreases. J Agric Food Chem. 2020; 68: 9205-9212.
- Gonçalves RFS, Martins JT, Abrunhosa L, Vicente AA, Pinheiro AC. Nanoemulsions for enhancement of curcumin bioavailability and their safety evaluation: Effect of emulsifier type. Nanomaterials. 2021; 11: 815.
- Chevalier L, Plourde M. Comparison of pharmacokinetics of omega-3 fatty acid supplements in monoacylglycerol or ethyl ester in humans: a randomized controlled trial. Eur J Clin Nutr. 2021; 75: 680-688.
- Moura ELR, dos Santos H, Celes APM, Bassani TB, Souza LC, et al. Effects of a Nutritional Formulation Containing Caprylic and Capric Acid, Phosphatidylserine, and Docosahexaenoic Acid in Streptozotocin-Lesioned Rats. J Alzheimer’s Dis Reports. 2020; 4: 353-363.
- Boison D. New insights into the mechanisms of the ketogenic diet. Curr Opin Neurol. 2017; 30: 187-192.
- Warren EC, Dooves S, Lugarà E, Damstra-Oddy J, Schaf J, et al. Decanoic acid inhibits mTORC1 activity independent of glucose and insulin signaling. Proc Natl Acad Sci U S A. 2020; 117: 23617-625.
- Ziętek M, Celewicz Z, Kikut J, Szczuko M. Implications of SCFAs on the parameters of the lipid and hepatic profile in pregnant women. Nutrients. 2021; 13: 1-13.
- Wang GQ, Bonkovsky HL, De Lemos A, Burczynski FJ. Recent insights into the biological functions of liver fatty acid binding protein 1. J Lipid Res. 2015; 56: 2238-2247.
- Gallardo D, Amills M, Quintanilla R, Pena RN. Mapping and tissue mRNA expression analysis of the pig solute carrier 27A (SLC27A) multigene family. Gene. 2013; 515: 220-223.
- Agame-Lagunes B, Grube-Pagola P, García-Varela R, AlexanderAguilera A, García HS. Effect of curcumin nanoemulsions stabilized with MAG and DAG-MCFAs in a fructose-induced hepatic steatosis rat model. Pharmaceutics. 2021; 13: 509.
- Esperón-Rojas AA, Baeza-Jiménez R, Cano-Sarmiento C, García HS. Structured Mono- and Diacylglycerols with a High Content of Medium Chain Fatty Acids. J Oleo Sci. 2017; 66: 991-996.
- Kawasaki T, Igarashi K, Koeda T, Sugimoto K, Nakagawa K, et al. Rats Fed Fructose-Enriched Diets Have Characteristics of Nonalcoholic Hepatic Steatosis. J Nutr. 2009; 139: 2067-2071.
- Kohli R, Kirby M, Xanthakos SA, Softic S, Feldstein AE, et al. HighFructose Medium-Chain-Trans-Fat Diet Induces Liver Fibrosis & Elevates Plasma Coenzyme Q9 in a Novel Murine Model of Obesity and NASH. Hepatology. 2010; 52: 934-44.
- Skenderian S, Park G, Jang C. Organismal fructose metabolism in health and non-alcoholic fatty liver disease. Biology (Basel). 2020; 9: 1-17.
- Spruss A, Kanuri G, Stahl C, Bischoff SC, Bergheim I. Metformin protects against the development of fructose-induced steatosis in mice: Role of the intestinal barrier function. Lab Investig. 2012; 92: 1020-1032.
- Gawrieh S, Chalasani N. Pharmacotherapy for Non-alcoholic Fatty Liver Disease. Physiol Behav. 2017; 176: 139-148.
- Zheng B, McClements DJ. Formulation of more efficacious curcumin delivery systems using colloid science: Enhanced solubility, stability, and bioavailability. Molecules. 2020; 25: 1-25.
- Miranda CA, Schönholzer TE, Klöppel E, Sinzato YK, Volpato GT, et al. Repercussions of low fructose-drinking water in male rats. An Acad Bras Cienc. 2019; 91: 1-10.
- Roeb E, Weiskirchen R. Fructose and Non-Alcoholic Steatohepatitis. Front Pharmacol. 2021; 12: 1-11.
- Maithilikarpagaselvi N, Sridhar MG, Swaminathan RP, Sripradha R, Badhe B. Curcumin inhibits hyperlipidemia and hepatic fat accumulation in high-fructose-fed male Wistar rats. Pharm Biol. 2016; 54: 2857-63.
- Lee EJ, Hwang JS, Kang ES, Lee SB, Hur J, et al. Nanoemulsions improve the efficacy of turmeric in palmitate- and high fat diet-induced cellular and animal models. Biomed Pharmacother. 2019; 110: 181-189.
- Nomura K, Yamanouchi T. The role of fructose-enriched diets in mechanisms of nonalcoholic fatty liver disease. J Nutr Biochem. 2012; 23: 203-208.
- Watanabe S, Tsujino S. Applications of Medium-Chain Triglycerides in Foods. Front Nutr. 2022; 9: 1-21.
- Oseini AM, Cole BK, Issa D, Feaver RE, Sanyal AJ. Translating scientific discovery: The need for preclinical models of nonalcoholic steatohepatitis. Hepatol Int. 2018; 12: 6-16.
- Liu Y, Zhang Y, Zhang X, Xu Q, Yang X, et al. Medium-chain fatty acids reduce serum cholesterol by regulating the metabolism of bile acid in C57BL/6J mice. Food Funct. 2017; 8: 291-298.
- Chien YJ, Chang CY, Wu MY, Chen CH, Horng YS, et al. Effects of curcumin on glycemic control and lipid profile in polycystic ovary syndrome: Systematic review with meta-analysis and trial sequential analysis. Nutrients. 2021; 13: 1-14.
- Kuszewski JC, Wong RHX, Wood LG, Howe PRC. Effects of fish oil and curcumin supplementation on cerebrovascular function in older adults: A randomized controlled trial. Nutr Metab Cardiovasc Dis. 2020; 30: 625-33.
- Thota RN, Acharya SH, Garg ML. Curcumin and/or omega-3 polyunsaturated fatty acids supplementation reduces insulin resistance and blood lipids in individuals with high risk of type 2 diabetes: A randomised controlled trial. Lipids Health Dis. 2019; 18: 1-11.
- El-Gizawy MM, Hosny EN, Mourad HH, Abd-El Razik AN. Curcumin nanoparticles ameliorate hepatotoxicity and nephrotoxicity induced by cisplatin in rats. Naunyn Schmiedebergs Arch Pharmacol. 2020; 393: 1941-1953.
- Qin S, Huang L, Gong J, Shen S, Huang J, et al. Efficacy and safety of turmeric and curcumin in lowering blood lipid levels in patients with cardiovascular risk factors: A meta-analysis of randomized controlled trials. Nutr J. 2017; 16: 1-10.
- Chen Q, Wang T, Li J, Wang S, Qiu F, et al. Effects of natural products on fructose-induced nonalcoholic fatty liver disease (NAFLD). Nutrients. 2017; 9: 12.
- Shen JD, Wei Y, Li YJ, Qiao JY, Li YC. Curcumin reverses the depressive-like behavior and insulin resistance induced by chronic mild stress. Metab Brain Dis. 2017; 32: 1163-1172.
- Atia MM, Abdel-Tawab HS, Mostafa AM, Mobarak SA. Nanocurcumin and curcumin prevent N, N’-methylenebisacrylamideinduced liver damage and promotion of hepatic cancer cell growth. Sci Rep. 2022; 12: 1-17.
- Hengist A, Koumanov F, Gonzalez JT. Fructose and metabolic health: governed by hepatic glycogen status? J Physiol. 2019; 597: 3573-3585.
- Monga SPS. Molecular Pathology of liver Diseases. 2011; 5.
- Zou L, Zheng B, Zhang R, Zhang Z, Liu W, et al. Influence of Lipid Phase Composition of Excipient Emulsions on Curcumin Solubility, Stability, and Bioaccessibility. Food Biophys. 2016; 11: 213-225.