
Journal of Gastroenterology Research and Practice
Case Series - Open Access, Volume 4
Plant-derived extracellular vesicles: A novel approach for gastrointestinal tract diseases
Naz Ünsal1; Nezih Başaran1; Ayşim Çisen Akyol2; Hatice Sönmez1; Alara Yalaza1; Merve Yıldırım Canpolat1*; Polen Koçak Denizci1*
1Cellestetix Genetic Consultancy R&D and Biotechnology Inc, Turkey.
2Yıldız Technical University, Bioenginerring Department, Turkey.
*Corresponding Author: Polen Koçak Denizci
Lecturer, Faculty of Engineering and Natural Sciences/
Biomedical Engineering, Istinye University, Turkey.
Cellestetix Genetic Consultant R&D and Biotechnology
Inc, Turkey.
Tel: +90 (532) 666 56 64;
Email: polen.kocak@istinye.edu.tr
Merve Yıldırım Canpolat
Cellestetix Genetic Consultancy R&D and Biotechnology
Inc, Turkey.
Tel: +90 (538) 421 61 21;
Email: merve@genestetix.com
Received : Jan 18, 2024
Accepted : Feb 07, 2024
Published : Feb 14, 2024
Archived : www.jjgastro.com
Copyright : © Yıldırım Canpolat, Koçak Denizci P (2024).
Abstract
Exosomes, nanovesicles with diameters ranging from 30 to 150 nm are crucial for intercellular communication. They are produced by mammalian and plant cells to transport bioactive components such as proteins, nucleic acids, metabolites, and lipids, which vary based on their origin. Although exosomes obtained from mammalian cells hold potential as therapeutic carriers, there are challenges in obtaining large quantities, and potential for triggering an immune response. Plant-derived extracellular vesicles (PDEVs) emerge as a potential solution, characterized by larger size and advantageous properties, including enhanced oral bioavailability, and resistance to degradation in high acidic conditions. Bioactive components of PDEVs contribute to various therapeutic properties, including anti-tumorigenic, anti-oxidative, anti-inflammatory effects, and the promotion of tissue regeneration. This review examines the formation, analysis, and new therapeutic possibilities of PDEVs in gastrointestinal tract (GIT) diseases, with a particular emphasis on their ability to regulate the immune system and maintain GI balance. It also discusses the future prospects and difficulties in developing nanotherapeutics for these diseases.
Citation: Ünsal N, Başaran N, Çisen Akyol A, Yıldırım Canpolat, Koçak Denizci P, et al. Plant-derived extracellular vesicles: A novel approach for gastrointestinal tract diseases. J Gastroenterol Res Pract. 2024; 4(2): 1183.
Introduction
Exosomes are small extracellular vesicles (EVs) that play a crucial role in intercellular communication within the body. These nanosized vesicles, ranging from 30 to 150 nanometers in diameter, are released by various cell types and carry a cargo of diverse molecules, including proteins, lipids, nucleic acids (such as RNA and DNA), and metabolites [1,2]. Exosomes are formed through the endocytic pathway, where intracellular vesicles are generated within multivesicular bodies (MVBs) and then released into the extracellular space when these MVBs fuse with the cell membrane [3]. This process allows exosomes to transport their molecular cargo to recipient cells, influencing various physiological and pathological processes.
A variety of sources have been identified as sources of EVs. For instance, they are produced by bacteria and are present in the bodily fluids of mammals, such as blood, urine, and plasma [4]. Exosomes obtained from mammalian cells show significant promise as therapeutic carriers. Nonetheless, the separation and effective utilization of these exosomes pose challenges in delivering numerous molecules to the target cells [5,7]. Aside from their difficulty in obtaining large quantities, they have the potential to trigger an immune response in the host [8]. Plantderived exosomes offer a potential solution to address these difficulties [9]. In recent years, there has been increased relevance to the potential functions of plant-derived extracellular vesicles (PDEVs) concerning health and disease. PDEVs are characterized by a larger particle size, ranging from 30 to 400 nm [10]. These nano-sized membrane vesicles are extracted from fresh plants and contain a mix of lipids, proteins, nucleic acids, and other small molecular components [11]. PDEVs offer distinct advantages when compared to exosomal nanoparticles derived from animal cells, including enhanced oral bioavailability and heightened resistance to degradation in acidic environments [12]. In contrast to exosomes derived from animal cells, PDEVs can be conveniently administered orally, rendering them more readily available and practical for therapeutic applications. In addition, they offer several benefits as vehicles for disease treatment compared to synthetic nanocarriers, displaying higher internalization rates, lower biological toxicity and immunogenicity, great targeting capacity and lower production costs for a large scale [13]. Also, they exhibit stability in the gastrointestinal tract and possess the capability to traverse the blood– brain barrier [14].
Presently, Plant-derived Extracellular Vesicles (PDEVs) have demonstrated beneficial immunoregulatory effects on gastrointestinal (GI) homeostasis. Inflammation is well recognized as a part of the innate immune response. If not controlled, it can develop into acute or chronic inflammatory illnesses, which can act as a primary cause for different chronic diseases including inflammatory bowel disease [15,16]. PDEVs have been identified as contributors to anti-inflammatory effects through the regulation of host immunological processes. Additionally, the miRNAs found in PDEVs play a role in mediating interkingdom communication between the gut microbiota and the host immune system, thereby contributing to the maintenance of a balanced state between immunity and gut microbiota [17,19]. Following internalization into recipient cells of the host, PDEVs activate various intracellular signaling pathways, modulating cellular responses and promoting tissue homeostasis [9,20].
PDEVs have been shown to have therapeutic effects on various digestive tract diseases, such as Inflammatory Bowel Disease (IBD), Ulcerative Colitis (UC), Crohn’s Disease (CD), Colitis Associated Cancer (CAC) and Colorectal Cancer [21]. PDEVs can modulate the immune system, regulate the gut microbiota, enhance the intestinal barrier function, and inhibit the proliferation and invasion of cancer cells [20,21]. In this review, we summarize the current knowledge on the biogenesis, characterization, and therapeutic potential of PDEVs for gastrointestinal tract (GIT) diseases.
Biogenesis of PDEVs (Plant Derived Extracellular Vesicles)
Plant-Derived Extracellular Vesicles (PDEVs) are made up of a complex web of mechanisms and activities. Although PDEVs have been successfully isolated and identified through research, the precise methods by which the biogenesis of these nanoscale membrane vesicles takes place are still unknown [22]. Three mechanisms have been hypothesized to release PDENs: the vacuolar pathway, the Exocyst Positive Organelle (EXPO) pathway, and the Multivesicular Body (MVB) pathway [23,24]. The Multivesicular Body (MVB) route, first identified in suspension culture cells of carrots in the 1960s, can be linked to the first pathway of PDEVs [25].
PDEV biogenesis usually starts with the cell’s perception of particular signals. Numerous times, pathogen infection, cellular stress, or other environmental variables can produce these signals. Furthermore, signals start the process of sending specific proteins and other cell components toward the MVB. The Multivesicular Body (MVB) pathway is one of these mechanisms that is particularly crucial to the biogenesis of PDENs. By meticulously forming membrane vesicles inside the cell and releasing their contents in a regulated manner outside of it, the MVB route offers a sophisticated mechanism [12]. Exosomes are produced by the endocytic pathway, in which intraluminal vesicles (ILVs) within Multivesicular Bodies (MVBs) are formed through membrane invasion that takes place during late endocytosis, a difficult process inside the cell [26]. An endosome that has been prematurely sorted is formed when the plasma membrane sprouts inward. This is the point at which a late-separating endosome enters the maturation process [12]. At this stage, the endosome undergoes a transition into Multivesicular Bodies (MVB) due to interaction between the late-sorted endosome and the trans-Golgi network [12]. Multivesicular bodies are a crucial step of cell evolution that emerges from the Trans-Golgi network (TGN)/Early Endosomes (EE) during the maturation process [27]. The vesicles that bud from the limiting membrane into the lumen are the source of distinct Intraluminal Vesicles (ILVs) of MVBs. This process is mediated by the protein complexes ESCRT-0, I, II, and III (endosomal sorting complex necessary for transport) [28]. ESCRT is made up of four complexes called ESCRT-0, -I, -II, and -III, which are made up of different proteins. Plants include TOM1-like (TOL) proteins, which bind ubiquitin and are necessary for endocytosis and vacuolar sorting of the auxin efflux transporter component PIN2, even though they lack conventional ESCRT-0 subunits [29]. Additionally, ESCRT-1 components recruit ESCRT-II and assemble widely occurring proteins. The assembly of ESCRT-III on the endosomal membrane for material sorting, concentration regulation, and vesicle formation is initiated by interactions between the Vps25 subunit of ESCRT-II and the Vps20/CHMP6 subunit of ESCRT-III [28]. After every cycle, ESCRT-III momentarily gathers on the endosomal membrane before separating into the cytoplasm [30]. MVB associates with lysosomes or the plasma membrane during the subsequent stage of biogenesis. When nanoparticles fuse with the plasma membrane, they are liberated from the cell [31]. The Rab GTPase family, which is present in the outer Multivesicular Body (MVB) membrane, and the soluble NSF binding receptor (SNARE), which is present after fusion, form the two protein complexes that regulate this cycle [32,33]. Enzymatic breakdown of the contents of Multivesicular Body (MVBs) is initiated by fusion with lysosomes [34]. This phase permits the cell to break down different proteins and other substances. One of the fundamental systems of the cell for cleaning and recycling, lysosomes help break down MVB content and facilitate the conversion of chemicals inside the cell into building blocks and energy [35]. This integration entails the lysosomal digestion of the exosomal contents and their subsequent fusing with the plasma membrane. The clearance, recycling, and release of chemicals within the cell are regulated by this mechanism. PDENs’ extracellular release facilitates the network of interactions and communication between these crucial cellular structures, which in turn supports a variety of biological functions [12].
The release of peroxisome-like organelles created inside the cell out of the cell by fusion with the plasma membrane is referred to as the “Exosome Positive Organelle” (EXPO) pathway in plant cells [36]. Because they include hydrolytic enzymes and defence proteins, vacuoles that react to infections by bacterial pathogens are able to fuse with the plasma membrane and release defence materials into the extracellular environment [37]. According to recent research, vesicles and central vacuoles are important for the production and release of PDENs during plant defence activities. In this regard, it was found that MVB vesicles are the source of the central vacuole in plant cells. This discovery represents a significant advancement in our knowledge of plants’ defensive mechanisms against infections [38].
Biological composition and function of PDEVs
Each plant has a different exosome composition, which affects bioactivity and functions like uptake, targeting, and gene expression modification. Exosomes comprise a combination of lipids, proteins, and nucleic acids, and the characteristics of these constituents can vary between different cell types [39]. The exosome surface is enriched with tetraspanins like CD63, CD81, and CD9, heat shock proteins such as HSP70, and various proteins involved in cell-cell communication, serving as key biomarkers for exosome characterization [40]. Both mammalian and plant exosomes contain nucleic acids, specifically mRNA and miRNAs, with mammalian exosomes typically containing around 100-300 miRNAs compared to fewer than 100 in plant-derived vesicles [41]. While mammalian exosomes exhibit a lipid composition rich in cholesterol but lower in phospholipids, plant-derived vesicles consist of up to 98% phospholipids, including plant-specific lipids like galactolipids [41]. Plant exosomes typically display a spherical structure with essential lipid bilayers [42]. Exosomes, owing to their natural properties and size, have the inherent ability to transport molecules such as drugs, chemotherapeutic agents, proteins, or siRNA to target cells [43]. In recent years, due to the capacity of exosomes derived from plants’ ability to create cross-kingdom communication through dietary absorption the plant-derived nanovesicles are starting to receive a lot of attention from researchers [44]. They have numerous therapeutic qualities, including anti-tumorigenic, anti-oxidative, and anti-inflammatory effects, because of their bioactive components [45].
Recent research, particularly Wang et al. has demonstrated that nanoparticles derived from grapefruit can effectively deliver specific molecules to cells [46]. Nanovesicles, akin to plant exosomes, play a significant role in drug distribution [39]. Modifying these molecules chemically enhances their ability to reach target tissues, minimizing biological distribution to non-target areas and subsequently increasing their immunogenicity. Several studies, including cytotoxicity assays, have been conducted to assess the toxicity of exosomes from edible plants [39]. For instance, research on exosomes from ginger, wheat, and grapefruit revealed that they do not exhibit toxic properties. Q. Wang et al. found grapefruit nanoparticles to be more stable than cationic liposomes, while M. Zhang et al. observed that ginger nanoparticles maintain stability in areas like the gut or stomach [46,47].
Exosome secretion and intake into the target cell
Exosomes may be involved in a wide range of biological processes and cellular communication channels, according to this variety [48]. An alternate pathway of the endocytic recycling process is represented by exosomes. An early endosome is created as the initial stage of the process. It buds inward from the plasma membrane and is then released into the cytoplasm of the cell. Subsequently, Intraluminal Vesicles (ILVs) sprout into the endosome by squeezing inward across the endosomal limiting membrane. In the meantime, ILVs extract material from the cytoplasm, which causes the endosomal membrane to form inside out and the endosomal content to be enriched. Intraluminal Vesicles (ILVs) congregate and compress inward, causing the endosome to transform into a Multivesicular Body (MVB). In order to maintain endosomal activities within the cell, the Multivesicular Body (MVB) typically merges with a lysosome. Because of this connection, lysosomal enzymes break down Intraluminal Vesicles (ILVs) that are housed within the MVB [49]. The fusion of MVBs with the plasma membrane is necessary for the last stage of exosome secretion. It is believed that a particular combination of soluble N-ethylmaleimide-sensitive factor binding protein receptors (SNAREs) is involved in this process. MVBresident vesicular SNAREs (v-SNAREs) engage with cell-resident target SNAREs (t-SNAREs) to start the fusion process with the plasma membrane. The release of exosomes from inside the cell to the outside of the cell occurs in a predictable manner as a result of this interaction [48]. Membrane fusion is caused by a SNARE complex, which is formed when a protein complex known as membrane bridging engages the plasma membrane [50]. Target SNAREs (t-SNAREs), located inside the cell, and vesicular SNAREs (v-SNAREs), localized in MVBs, interact with the plasma membrane in this process by forming a particular combination [51]. Exosomes are released from within the cell and onto the exterior of the cell as a result of this interaction, which starts the membrane fusion process [49]. The membrane-bridging SNARE complex is involved in cellular communication and helps to maintain a certain, controlled oscillation rhythm [49]. Research has shown that a few different factors influence the secretion of exosomes [49]. It was demonstrated by Fader and colleagues that the TI-VAMP/VAMP7 v-SNARE complex, which is present in the erythroleukemia cell line, actively contributes to the fusion of Multivesicular Bodies (MVBs) with the plasma membrane [51].
Exosome release is triggered by cell activation brought on by the cross-linking of receptors such as CD3 on T cells or IgE on mast cells [52]. Also, exosome membrane proteins can communicate with target cells through a variety of methods [49]. First, exosome membrane proteins can directly connect with target cell receptors to initiate intracellular signaling. A secondary mechanism involves the cleavage of exosome membrane proteins by proteases, resulting in soluble fragments that have the ability to attach themselves to cell surface receptors and function as soluble ligands. Third, target cells have the ability to ingest exosomes and release the material therein, causing subsequent actions to occur in recipient cells. Exosomes are dynamic entities that can connect with target cells in a variety of ways, as evidenced by these various modes of contact [48]. When exosomal membrane ligands attach to target cell receptors, a variety of interactions can be started. The ligand-receptor contact is one of these interactions that causes the exosomal membrane to fuse with the cell’s plasma membrane, releasing the cargo into the cytosol. Simultaneously, a downstream signaling cascade in the cell is started by receptor-ligand interaction, and this signaling cascade may help regulate the target cell’s biological responses. The exosome finally undergoes endocytosis, which is followed by fusion of the exosome with the endosome membrane, releasing the cargo into the cytosol [49].
Therapeutic effects of PDEVs on digestive tract diseases
There has been an increase in the use of edible plants for the isolation of exosomes in different areas of research as well as digestive tract disorders over the last few years (Figure1). A prevalent rationale for employing edible plant-derived exosomes in therapy lies in their ability to exert regulatory effects on a diverse array of anti-inflammatory cytokines within the intestine, coupled with their demonstrated antitumor activities and regenerative effects. Investigated plants so far include grapes, ginger, grapefruit, broccoli, turmeric, tea and mulberry bark.
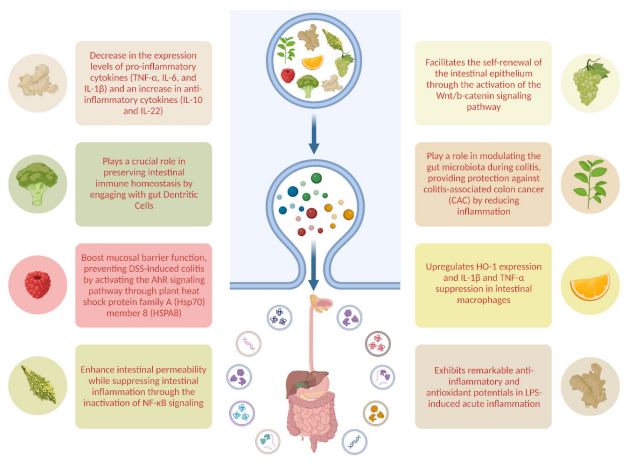
Grape Derived Extracellular Vesicles (GDEVs)
Studies directed by Ju et al. indicate that GDENs exhibit a diverse protein composition, encompassing heat shock protein 70 (HSP70), aquaporin, and anti-inflammatory lipids [53]. Impressively, GDENs demonstrate the ability to diffuse through the intestinal mucus barrier and undergo internalization by intestinal stem cells [53]. This process stimulates intestinal stem cell proliferation, facilitating the self-renewal of the intestinal epithelium through the activation of the Wnt/b-catenin signaling pathway [53]. This, in turn, prevents Dextran Sulfate Sodium (DSS)-induced colitis in mice, swiftly restoring the entire length of the intestinal architecture. Additionally, another study showed that grape derived extracellular vesicles, when administered orally, demonstrated resilience against degradation in saliva, the acidic stomach environment, and the potent proteolytic enzymes in the intestinal tract [54]. Recent findings indicate that these edible plant-based nanoparticles can effectively reach the intestine through oral delivery, potentially being absorbed by intestinal cells to perform functions like promoting intestinal regeneration [54]. This suggests that plant-derived edible nanoparticles, with a natural affinity for colonic tissues and anti-inflammatory properties, could serve as an innovative, safe, and scalable delivery system for addressing digestive tract diseases like Ulcerative Colitis (UC) and Inflammatory Bowel Disease (IBD) [53,54].
Ginger Derived Extracellular Vesicles (GDEVs)
Ginger, derived from the rhizome of Zingiber officinale, stands as one of the most extensively utilized natural products [55]. Therapeutic applications of ginger extend to the treatment of nausea and various digestive tract issues such as colic, flatulence, diarrhea, and dyspepsia [56]. Historically, ginger has been employed to regulate intestinal microflora and maintain intestinal balance among individuals with gastrointestinal disorders [57]. Notably, researchers indicate that ginger and its key components, such as 6-gingerol and 6-shogaol, show antioxidative, anti-inflammatory, and anti-cancer properties [58].
The oral administration of GDEVs demonstrated a notable reduction in both acute and chronic inflammation, exhibiting efficacy in mitigating inflammatory bowel diseases, colitis, and colitis-associated cancer [59,60]. Moreover, GDEVs exhibited a capacity to facilitate the healing of the intestinal mucosa, indicative of their potential in preventing chronic colitis and inhibiting tumor development [61]. This effect was associated with a decrease in the expression levels of pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β) and an increase in anti-inflammatory cytokines (IL-10 and IL-22) [42,62]. These may indicate that EVs inhibit factors that damage the intestine and encourage intestine healing factors. Furthermore, GDEVs display a selective uptake by Lactobacillaceae and target specific genes of Lactobacillus rhamnosus (LGG) [12]. This interaction plays a role in changing the composition of gut microbiota and triggering the expression of IL-22, leading to the restoration of intestinal barrier function and the alleviation of colitis [12]. Notably, ginger extract itself exhibits the capacity to modulate dysbiosis in the intestinal microbiota, enhances intestinal barrier function, and suppresses the inflammatory response in Dextran Sulfate Sodium (DSS)- induced colitis [59,63,64].
In models of chemically induced colorectal cancer, the oral administration of nanoparticles derived from ginger resulted in a substantial reduction in tumor size. This reduction can be attributed to the inhibition of apoptosis, the proliferation of intestinal epithelial cells, and the decrease in pro-inflammatory cytokines. The treatment with ginger exosomes was observed to potentially upregulate 14 proteins and downregulate 3 proteins, including noteworthy molecular targets such as PKG and transgelin, which have previously been associated with colon cancer [26]. This signifies the promising impact of ginger-derived nanoparticles in modulating key molecular pathways implicated in colorectal cancer development.
Grapefruit Derived Extracellular Vesicles (GFDEVs)
Grapefruit-derived extracellular vesicles (GFDEVs) are rich in phosphatidylethanolamine and phosphatidylcholine, lipids known for their antioxidant, anti-inflammatory, and anti-colitic properties [65]. The primary flavonoid in grapefruit, naringin, undergoes hydrolysis by intestinal microflora to produce the active compound naringenin [65]. Naringenin possesses diverse pharmacological benefits, like anticancer, anti-inflammatory, and antioxidant effects. The intake of GFDEVs by intestinal macrophages stimulates the expression of the HO-1 and IL-10, while inhibiting the production of proinflammatory cytokines IL-1β and TNF-α [65,66]. Studies by Wang et al. demonstrated that Grapefruit-Derived Nanovesicles (GDNs) selectively target intestinal macrophages, alleviating DSS-induced mouse colitis [65]. These anti-inflammatory actions of GDNs are caused by upregulating HO-1 expression and IL-1β and TNF-α suppression in intestinal macrophages. Also, regarding the authors, GDNs were found to be biocompatible, biodegradable, and stable across varying pH levels, suggesting their potential as an innovative oral drug delivery system. Incorporating the anti-inflammatory drug Methotrexate (MTX) into GDNs resulted in reduced toxicity compared to free MTX, displaying significantly enhanced therapeutic effects against DSS-induced colitis in mice. These findings highlight the immune-modulating potential of GDNs in the intestine, offering promise as an oral delivery system for small molecule drugs targeting inflammatory responses in human diseases [65]. Hence, the substantial potential of grapefruit-derived extracellular vehicles (EVs) lies in their ability to modulate intestinal immune responses and potentially serve as a treatment for colitis in the future [21].
Broccoli Derived Extracellular Vesicles (BDEVs)
Boasting a wealth of vitamins, carotenoids, dietary fiber, minerals, and flavonoids, broccoli (Brassica oleracea var. italica) stands as a prevalent cruciferous vegetable globally [67]. With its copious nutrients and phytochemicals, broccoli proves advantageous in modifying the gut microbiome population, thereby contributing to the prevention and treatment of specific diseases [68,69]. BDEVs that have elements characterized by low immunogenicity, high gastrointestinal stability, the capacity to overcome biological barriers, cost-effectiveness, and the ability for mass production—such as proteins, lipids, miRNAs, and other active components—play a pivotal role in the prevention of intestinal diseases like colitis [70,71].
Adenosine Monophosphate-Activated Protein Kinase (AMPK) is gaining prominence as a crucial enzyme and pathway involved in regulating immune homeostatic networks [72]. Expressed in diverse immune cell types like macrophages, lymphocytes, neutrophils, and Dendritic Cells (DCs), AMPK governs various immune functions, including cytokine production, chemotaxis, cytotoxicity, apoptosis, and proliferation [73,74]. A study delved into the effects of Broccoli-Derived Extracellular Vesicles (BDEVs) on the immune system, revealing that broccoli nanoparticles primarily target Dendritic Cells (DCs) [75]. This targeting leads to the induction of tolerogenic DCs, demonstrating a preventive effect against mouse colitis [75]. Moreover, the research highlighted the pivotal role of DC AMPK in the lipid-mediated induction of tolerogenic DCs by Broccoli-Derived nanoparticles (BDEVs) [75]. Additionally, it underscored the contribution of Sulforaphane (SFN), carried by BDNs, in preventing mouse colitis through AMPK-mediated induction of tolerogenic DCs [75]. Collectively, these findings suggest that nanoparticles from broccoli play a crucial role in preserving intestinal immune homeostasis by engaging with gut DCs [75]. This understanding could contribute to the development of healthier dietary recommendations aimed at preventing both acute and chronic inflammatory diseases such as IBD [76,77]. Importantly, this approach seeks to minimize potential adverse reactions associated with the metabolic energy effects of AMPK. In another study, BDEVs were shown to alleviate LOP-induced constipation in mice by influencing gastrointestinal hormones, gut microbial structure, and intestinal metabolites [78]. These research findings underscore the promising potential of developing BDEVs as safe and effective nano-therapies for the treatment of constipation [78].
Turmeric Derived Extracellular Vesicles (TDEVs)
Curcumin, a natural hydrophobic polyphenol extraction from turmeric’s rhizomes (Curcuma longa), is renowned for its diverse pharmacological effects [79]. Promising outcomes from preclinical investigations highlight its antioxidant, anti-inflammatory, antitumor, immunomodulatory, wound healing, antiproliferative, and antimicrobial properties [80]. Due to its innocuous nature, affordability, and availability, both in vitro and in vivo studies suggest curcumin’s potential as a therapeutic agent against chronic conditions such as inflammatory bowel disease, arthritis, pancreatitis, chronic anterior uveitis, and cancers [80,81]. Notably, oral administration ensures an active curcumin level in the gastrointestinal tract, positioning it as a promising candidate for diseases affecting this anatomical site [82].
Research conducted by Liu et al. underscores the rich lipid and protein content of TDEVs, contributing to their impressive anti-inflammatory and antioxidant attributes [83]. Their studies also demonstrate that TDEVs exhibit remarkable antiinflammatory and antioxidant potentials in LPS-induced acute inflammation. In mouse models of colitis, orally administered TDEVs selectively accumulate in the inflamed colon, primarily internalizing within colonic epithelial cells and macrophages [83]. TDEVs alleviate DSS-induced colitis in mice by mitigating damaging factors and enhancing protective elements, thereby promoting inflammation resolution in the colon [83]. The inactivation of the NF-κB pathway by TDEVs provides a potential explanation for their protective effects against colitis [83]. In conclusion, the study results collectively propose TDEVs as a novel therapeutic approach for colitis, offering functions in both inflammation prevention and resolution.
Tea Leaf Derived Extracellular Vesicles (TLDEVs)
Tea stands out as one of the globally popular and extensively consumed beverages. Its appeal is not only due to its flavor but also stems from its health-enhancing attributes, including antioxidant, antitumor, and hypolipidemic properties [84]. These benefits arise from the presence of beneficial compounds like polyphenols, flavones, lipids, and polysaccharides in tea leaves [85,86]. A prime example is the utilization of tea leaf extracts rich in EpiGallocatechin-3-Gallate (EGCG) to alleviate colitis by eliminating Reactive Oxygen Species (ROS), mitigating mast cell activation, and curtailing inflammatory responses [87]. Another noteworthy polyphenol found in tea leaves, Epicatechin Gallate (ECG), not only directly combats ROS but also elevates the levels of heme oxygenase-1 (HO-1) and glutathione (GSH), exhibiting robust antioxidant activity [88]. Furthermore, research conducted by Zu et al. indicates that polysaccharides from tea leaves play a role in modulating the gut microbiota during colitis, providing protection against Colitis-Associated Colon Cancer (CAC) by reducing inflammation [89]. In the scenario of mouse colitis, macrophages selectively internalize TLDENs following oral administration. Moreover, oral administration of tea leafderived nanoparticles effectively suppresses inflammatory bowel responses, restores compromised colonic barriers, and enhances the diversity and overall abundance of the gut microbiota in mouse colitis [84].
Mulberry Bark Derived Extracellular Vesicles (MBDEVs)
A recent study revealed that mulberry bark-derived ELNs (MBELNs) can boost mucosal barrier function, preventing DSS-induced colitis by activating the AhR signaling pathway through plant heat shock protein family A (Hsp70) member 8 (HSPA8) [90]. AhR pathway activation in intestinal epithelial cells prompts the expression of COP9 signalosome subunit 8 (COPS8), a key regulator of proteasome degradation and neddylation [90]. Neddylation is a post-translational modification analogous to ubiquitination in which the ubiquitin-like protein neural precursor cell expressed developmentally downregulated protein 8 (NEDD8) attaches to its substrate proteins [91]. The COP9 signalosome suppresses Cullin–RING E3 ligase activity by enhancing NEDD8–Cullin 1 conjugate cleavage, leading to increased Cullin 1 de-neddylation following MBELN treatment [92]. Induction of COPS8/COP9 through MBEAN-mediated AhR pathway activation results in the production of Antimicrobial Peptides (AMPs), which hinder intestinal inflammation and prevent DSS-induced colitis [90]. AMPs combat infections through direct antimicrobial activity and/or by influencing the host immune response [90]. Additionally, AMPs exhibit anti-inflammatory, antitumor, immunoregulatory, and various biological effects. They selectively inhibit pro-inflammatory responses, such as TNF and ROS production, by inhibiting toll-like receptor 4 (TLR4)-activated TLR signaling pathway and downstream pathways like NF-κB and MAPK [93]. The immunomodulatory roles of AMPs encompass recruitment of leukocytes to infection sites, modulation of neutrophil responses, enhancement of phagocytosis, differentiation of dendritic cells, and recruitment and polarization of T cells [94]. Furthermore, the crosstalk between COPS8/COP9 and the proteasome degradation pathway modulates MBELN-mediated AhR signaling homeostasis [90].
Momordica Charantia-Derived Extracellular Vesicles (MCDEVs)
Bitter melon, scientifically known as Momordica Charantia (MC), is a rich source of carbohydrates, crude protein, dietary fiber, and vitamin C [95]. Extensive research has highlighted its diverse health benefits, including anti-oxidative, anti-inflammatory, anti-cancer, and hypoglycemic effects, positioning it as a potential remedy for inflammation and cancer [96,97]. Semiz et al.’s studies revealed that MC-homogenized jam effectively reduced inflammation in the colon tissue of rats [98]. Furthermore, MC polysaccharides were found to enhance intestinal permeability while suppressing intestinal inflammation through the inactivation of NF-κB signaling [98]. Notably, proteins derived from MC seeds demonstrated antioxidant properties, regulating oxidative stress and mitigating inflammatory factors in vitro, though exhibiting contrasting effects in vivo, potentially attributed to the method of intraperitoneal administration [98]. Researchers have investigated the lipid composition and proteomics analysis of MC extracellular vesicles (MCEVs), revealing their promising antioxidant capabilities [99]. This suggests their potential as a natural therapeutic agent for conditions such as obesity, diabetes, inflammation, and cancer [99]. Subsequent experiments involving mice with Ulcerative Colitis (UC) demonstrated that MCEVs could safeguard the colonic mucosa by modulating oxidation and inflammation indexes in the bloodstream, alleviating symptoms of colonic ulceration [99]. Concurrently, MCEVs significantly suppressed levels of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) and elevated the level of the anti-inflammatory cytokine IL-10 in mouse serum [99]. These findings underscore the crucial role of antioxidant protection in MCEVs’ impact on UC, suggesting their potential as a therapeutic drug or carrier for various oxidative stress-induced diseases [99].
Conclusion
Plant derived extracellular vesicles have attracted quite a lot of attention as a trendy study recently. Thanks to its advantages such as low toxicity, low production cost, low immunogenicity and high stability of plant -derived extracellular vesicles, promising studies are revealed in the field of medicine and health. Since the content and surface markers of plant based extracellular vesicles are not fully known, more studies are needed in these topics. However, in the studies conducted so far, plant-derived extracellular vesicles are seen as high potential for use and application in the field of biotechnology and medicine. Due to their benefits, plant based extracellular vesicles deserve more studies for revealing their unknown potential and existing therapeutic effects.
References
- Exosome biogenesis, bioactivities and functions as new delivery systems of natural compounds - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/29248680/.
- Full article: Plant-Derived Exosome-Like Nanovesicles: Current Progress and Prospects. Accessed. 2023. [Online]. Available: https://www.tandfonline.com/doi/full/10.2147/IJN.S420748.
- Hybrid exosomes, exosome-like nanovesicles and engineered exosomes for therapeutic applications - ScienceDirect. Accessed. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/abs/pii/S0168365922008422.
- LM Doyle , MZ. Wang, Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. 20198; 7: 727. doi: 10.3390/cells8070727.
- J Chen. Review on Strategies and Technologies for Exosome Isolation and Purification, Front Bioeng Biotechnol. 2021; 9: 811971. doi: 10.3389/fbioe.2021.811971.
- D Yang. Progress, opportunity, and perspective on exosome isolation - efforts for efficient exosome-based theranostics, Theranostics. 2020; 10(8): 3684. doi: 10.7150/thno.41580.
- E Tzng, N Bayardo, and PC Yang, Current challenges surrounding exosome treatments, Extracellular Vesicle. 2023; 2:100023. doi: 10.1016/j.vesic.2023.100023.
- Pharmaceutics | Free Full-Text | Plant-Derived Exosomes as a Drug-Delivery Approach for the Treatment of Inflammatory Bowel Disease and Colitis-Associated Cancer. Accessed. 2023. [Online]. Available: https://www.mdpi.com/1999-4923/14/4/822#B10-pharmaceutics-14-00822.
- Plant Exosome-like Nanovesicles: Emerging Therapeutics and Drug Delivery Nanoplatforms: Molecular Therapy. Accessed. 2023. [Online]. Available: https://www.cell.com/molecular-therapy-family/molecular-therapy/fulltext/S1525-0016(20)30656 0?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS1525001620306560%3Fshowall%3Dtrue.
- Current understanding of plant-derived exosome-like nanoparticles in regulating the inflammatory response and immune system microenvironment - ScienceDirect. Accessed. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S1043661823000890.
- L Yu, Z Deng, L Liu, W Zhang, and C Wang, Plant-Derived Nanovesicles: A Novel Form of Nanomedicine, Front Bioeng Biotechnol. 2020; 8: 584391. doi: 10.3389/fbioe.2020.584391.
- D Li. Plant-derived exosomal nanoparticles: potential therapeutic for inflammatory bowel disease, Nanoscale Adv. . 2023; 5(14): 3575-3588. doi: 10.1039/D3NA00093A.
- E Woith, G Fuhrmann, and MF Melzig, Extracellular VesiclesConnecting Kingdoms, Int J Mol Sci. 2019; 20(22): 5695. doi: 10.3390/ijms20225695.
- Biological properties and therapeutic effects of plant-derived nanovesicles - PMC. Accessed. 2023. [Online]. Available: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7718644/.
- Plant-derived extracellular vesicles -a novel clinical antiinflammatory drug carrier worthy of investigation - ScienceDirect. Accessed. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/pii/S075333222301702X?via%3Dihub.
- 1. Z. Xu et al., Plant-derived extracellular vesicles (PDEVs) in nanomedicine for human disease and therapeutic modalities, J Nanobiotechnology. 2023; 21:114. doi: 10.1186/s12951-023-01858-7.
- K. SONOYAMA and F. OHSAKA, Role of microRNAs in the crosstalk between the gut microbiota and intestinal immune system, Biosci Microbiota Food Health. 2023; 42(4); 222228. doi: 10.12938/bmfh.2023-027.
- E Díez-Sainz, S Lorente-Cebrián, P Aranaz, JI Riezu-Boj, JA Martínez, and FI Milagro, Potential Mechanisms Linking FoodDerived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health, Front Nutr.2021; 8: 586564. doi: 10.3389/fnut.2021.586564.
- İ Özkan et al. Garlic (Allium sativum)-derived SEVs inhibit cancer cell proliferation and induce caspase mediated apoptosis, Sci Rep. 2021; 11: 14773. doi: 10.1038/s41598-021-93876-4.
- M Zhang, E Viennois, C Xu, and D Merlin, Plant derived edible nanoparticles as a new therapeutic approach against diseases, Tissue Barriers. 2016; 4(2): 1134415. doi: 10.1080/21688370.2015.1134415.
- C Yang et al. Edible plant-derived extracellular vesicles serve as promising therapeutic systems, Nano TransMed. 2023; 2(2):100004. doi: 10.1016/j.ntm.2023.100004.
- Y Cui, J Gao, Y He, and L Jiang, Plant extracellular vesicles, Protoplasma. 2023; 257(1): 3-12. X, doi: 10.1007/s00709-019-01435-6.
- M Cong et al. Technology insight: Plant-derived vesicles-How far from the clinical biotherapeutics and therapeutic drug carriers?, Adv Drug Deliv Rev. 2022; 182: 114108. doi: 10.1016/j.addr.2021.114108.
- J Kim, S Li, S Zhang, and J Wang, Plant-derived exosome-like nanoparticles and their therapeutic activities, Asian J Pharm Sci. 2022; 17(1): 53-69. doi: 10.1016/j.ajps.2021.05.006.
- Ultrastructural changes during growth and embryogenesis in carrot cell cultures - ScienceDirect. Accessed. 2023. [Online].Available: https://www.sciencedirect.com/science/article/abs/pii/S002253206780128X.
- D Subha, K Harshnii, KG Madhikiruba, M Nandhini, and KS Tamilselvi, Plant derived exosome- like Nanovesicles: an updated overview, Plant Nano Biology. 2023; 3: 100022. doi: 10.1016/j.plana.2022.100022.
- Y Cui, J Shen, C Gao, X Zhuang, J Wang, and L Jiang, Biogenesis of Plant Prevacuolar Multivesicular Bodies, Mol Plant. 2016; 9(6): 774-786. doi: 10.1016/j.molp.2016.01.011.
- Molecular mechanism of multivesicular body biogenesis by ESCRT complexes | Nature. Accessed. 2023. [Online]. Available: https://www.nature.com/articles/nature08849.
- B Korbei. Arabidopsis TOL Proteins Act as Gatekeepers for Vacuolar Sorting of PIN2 Plasma Membrane Protein, Current Biology. 2013; 23(24): 2500-2505. doi: 10.1016/j.cub.2013.10.036.
- Biogenesis and Function of Multivesicular Bodies - PMC. Accessed. 2023. [Online]. Available: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2911632/.
- Classification, functions, and clinical relevance of extracellular vesicles - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/22722893/.
- SNARE-protein-mediated disease resistance at the plant cell wall | Nature. Accessed. 2023. [Online]. Available: https://www.nature.com/articles/nature02076.
- N Inada. Modulation of Plant RAB GTPase-Mediated Membrane Trafficking Pathway at the Interface Between Plants and Obligate Biotrophic Pathogens, Plant Cell Physiol. 2016; 57(9): 1854-1864. doi: 10.1093/pcp/pcw107.
- The deubiquitinating enzyme AMSH1 and the ESCRT-III subunit VPS2.1 are required for autophagic degradation in Arabidopsis - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/23800962/.
- Applied Sciences | Free Full-Text | Plant Exosomal Vesicles: Perspective Information Nanocarriers in Biomedicine. Accessed. 2023. [Online]. Available: https://www.mdpi.com/2076-3417/12/16/8262
- EXPO, an exocyst-positive organelle distinct from multivesicular endosomes and autophagosomes, mediates cytosol to cell wall exocytosis in Arabidopsis and tobacco cells - PubMed. Accessed: Dec. 16, 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/21193573/.
- Plant extracellular vesicles: Trojan horses of cross-kingdom warfare - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/34485834/.
- Plant-Derived Nano and Microvesicles for Human Health and Therapeutic Potential in Nanomedicine - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/33917448/.
- Polen Koçak, Edible plant-derived exosomes and their therapeutic applicatons, J Biomed Imag Bioeng. 2020; 4: 1.
- MP Caby, D Lankar, C Vincendeau-Scherrer, G Raposo, and C Bonnerot, Exosomal-like vesicles are present in human blood plasma, Int Immunol. 2005; 17(7): 879-887. doi: 10.1093/intimm/dxh267.
- Exosomes in cancer: small particle, big player - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/26156517/.
- M Zhang. Edible ginger-derived nanoparticles: A novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer, Biomaterials. 2016; 101: 321-340. doi: 10.1016/j.biomaterials.2016.06.018.
- M. Zhang et al. Edible Ginger-derived Nano-lipids Loaded with Doxorubicin as a Novel Drug-delivery Approach for Colon Cancer Therapy, Mol Ther. 2016; 24(10): 1783-1796. doi: 10.1038/mt.2016.159.
- X Ou. Novel plant-derived exosome-like nanovesicles from Catharanthus roseus: preparation, characterization, and immunostimulatory effect via TNF-α/NF-κB/PU.1 axis, Journal of Nanobiotechnology. 2023; 21(1): 160. doi: 10.1186/s12951-023-01919-x.
- A Li et al. Plant-derived nanovesicles: Further exploration of biomedical function and application potential, Acta Pharmaceutica Sinica B. 2023; 13(8): 3300-3320. doi: 10.1016/j.apsb.2022.12.022.
- Q Wang et al. Grapefruit-Derived Nanovectors Use an Activated Leukocyte Trafficking Pathway to Deliver Therapeutic Agents to Inflammatory Tumor Sites, Cancer Res. 2015; 75(12): 2520-2529. doi: 10.1158/0008-5472.CAN-14-3095.
- Advances and Challenges of Liposome Assisted Drug Delivery - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/26648870/.
- L. Urbanelli et al., Signaling Pathways in Exosomes Biogenesis, Secretion and Fate. 2013; 4(2): 2. doi: 10.3390/genes4020152.
- Stem cell-derived exosomes: a novel vector for tissue repair and diabetic therapy in: Journal of Molecular Endocrinology. 2017; 59: 4. Available: https://jme.bioscientifica.com/view/journals/jme/59/4/JME-17-0080.xml.
- M Chaineau, L Danglot, and T Galli, Multiple roles of the vesicular-SNARE TI-VAMP in post-Golgi and endosomal trafficking, FEBS Lett. 2009; 583(23): 3817-3826. doi: 10.1016/j.febslet.2009.10.026.
- TI-VAMP/VAMP7 and VAMP3/cellubrevin: two v-SNARE proteins involved in specific steps of the autophagy/multivesicular body pathways - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/19781582/.
- E.-M. Krämer-Albers et al., Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: Trophic support for axons?, PROTEOMICS – Clinical Applications. 2007; 1(11): 1446-146. doi: 10.1002/prca.200700522.
- S Ju. Grape Exosome-like Nanoparticles Induce Intestinal Stem Cells and Protect Mice From DSS-Induced Colitis, Mol Ther. 2013; 21(7): 1345-1357. doi: 10.1038/mt.2013.64.
- M Rahimi Ghiasi, E Rahimi, Z Amirkhani, and R Salehi, Leucinerich Repeat-containing G-protein Coupled Receptor 5 Gene Overexpression of the Rat Small Intestinal Progenitor Cells in Response to Orally Administered Grape Exosome-like Nanovesicles, Adv Biomed Res. 2018; 7: 125. doi: 10.4103/abr.abr_114_18.
- K. Srinivasan, Ginger rhizomes (Zingiber officinale): A spice with multiple health beneficial potentials, PharmaNutrition. 2017; 5(1): 18-28. doi: 10.1016/j.phanu.2017.01.001.
- D Fitriyanti and R Sulung, Effectiveness of ginger to overcome nausea and vomiting caused by chemotherapy in breast cancer patients, Can Oncol Nurs J. 2020; 30(1): 3-5. doi: 10.5737/2368807630135.
- M Nikkhah Bodagh, I Maleki, and A Hekmatdoost, Ginger in gastrointestinal disorders: A systematic review of clinical trials, Food Sci Nutr. 2018; 7(1): 96-108. doi: 10.1002/fsn3.807.
- H Zhu and W. He, Ginger: a representative material of herbderived exosome-like nanoparticles, Frontiers in Nutrition. 2023; 10. Available: https://www.frontiersin.org/articles/10.3389/fnut.2023.1223349.
- M Zhang, JF Collins, and D Merlin, Do ginger-derived nanoparticles represent an attractive treatment strategy for inflammatory bowel diseases?, Nanomedicine (Lond). 2016; 11(23): 3035-3037. doi: 10.2217/nnm-2016-0353.
- M Zhang, X Wang, MK Han, JF Collins, and D Merlin, Oral administration of ginger-derived nanolipids loaded with siRNA as a novel approach for efficient siRNA drug delivery to treat ulcerative colitis, Nanomedicine (Lond). 2017; 12(16): 1927-1943. doi: 10.2217/nnm-2017-0196.
- AC Brown, C Shah, J Liu, JT H Pham, JG Zhang, and MR Jadus, Gingers (Zingiber officinale Roscoe) inhibition of rat colonic adenocarcinoma cells proliferation and angiogenesis in vitro, Phytother Res. 2009; 23(5): 640-645. doi: 10.1002/ptr.2677.
- L Yin. Characterization of the MicroRNA Profile of Ginger Exosome-like Nanoparticles and Their Anti-Inflammatory Effects in Intestinal Caco-2 Cells, J. Agric. Food Chem. 2022; 70(15): 4725-4734. doi: 10.1021/acs.jafc.1c07306.
- S Guo et al. Ginger Alleviates DSS-Induced Ulcerative Colitis Severity by Improving the Diversity and Function of Gut Microbiota, Front Pharmacol. 2021; 12: 632569. doi: 10.3389/fphar.2021.632569.
- X Zhou et al. Ginger Extract Decreases Susceptibility to Dextran Sulfate Sodium-Induced Colitis in Mice Following Early Antibiotic Exposure. 2022; 8: 755969. doi: 10.3389/fmed.2021.755969.
- B Wang. Targeted drug delivery to intestinal macrophages by bioactive nanovesicles released from grapefruit, Mol Ther. 2014; 22(3): 522-534. doi: 10.1038/mt.2013.190.
- J Mu et al. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles, Mol Nutr Food Res. 2014; 58(7): 1561-1573. doi: 10.1002/mnfr.201300729.
- TE Tallei, BJ Kepel, HIS Wungouw, F Nurkolis, AA Adam, and Fatimawali, A comprehensive review on the antioxidant activities and health benefits of microgreens: current insights and future perspectives, International Journal of Food Science & Technology, vol. n/a, no. n/a, doi: 10.1111/ijfs.16805.
- G Zandani, S Anavi-Cohen, N Sela, A Nyska, and Z Madar, Broccoli consumption attenuates inflammation and modulates gut microbiome composition and gut integrity-related factors in mice fed with a high-fat high-cholesterol diet, Food & Nutrition Research. 2021. doi: 10.29219/fnr.v65.7631.
- A Yanaka, Daily intake of broccoli sprouts normalizes bowel habits in human healthy subjects, Journal of Clinical Biochemistry and Nutrition. 2018; 62(1): 75-82. doi: 10.3164/jcbn.17-42.
- L del Pozo-Acebo. Therapeutic potential of broccoli-derived extracellular vesicles as nanocarriers of exogenous miRNAs, Pharmacological Research. 2022; 185: 106472. doi: 10.1016/j.phrs.2022.106472.
- Molecules | Free Full-Text | Preparation, Characterization, and In Vitro Anticancer Activity Evaluation of Broccoli-Derived Extracellular Vesicle-Coated Astaxanthin Nanoparticles. Accessed. 2023. [Online]. Available: https://www.mdpi.com/1420-3049/27/12/3955.
- L Antonioli et al. The AMPK enzyme-complex: from the regulation of cellular energy homeostasis to a possible new molecular target in the management of chronic inflammatory disorders, Expert Opinion on Therapeutic Targets. 2016; 20(2): 179-191. doi: 10.1517/14728222.2016.1086752.
- SB Vasamsetti, S Karnewar, AK Kanugula, AR Thatipalli, JM Kumar, and S Kotamraju, Metformin Inhibits Monocyte-to-Macrophage Differentiation via AMPK-Mediated Inhibition of STAT3 Activation: Potential Role in Atherosclerosis, Diabetes, vol. 2014; 64(6): 2028-2041. doi: 10.2337/db14-1225.
- RR Ramiscal et al. Attenuation of AMPK signaling by ROQUIN promotes T follicular helper cell formation. eLife. 2015; 4: 08698. doi: 10.7554/eLife.08698.
- Broccoli-Derived Nanoparticle Inhibits Mouse Colitis by Activating Dendritic Cell AMP-Activated Protein Kinase: Molecular Therapy. Accessed. 2023. [Online]. Available: https://www.cell.com/molecular-therapy-family/molecular-therapy/fulltext/S1525-0016(17)30064-3?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS1525001617300643%3Fshowall%3Dtrue.
- G Garcin et al. AMPK/HuR-Driven IL-20 Post-Transcriptional Regulation in Psoriatic Skin, Journal of Investigative Dermatology. 2015; 135(11): 2732-2741. doi: 10.1038/jid.2015.282.
- Q Cao et al. Myeloid Deletion of α1AMPK Exacerbates Atherosclerosis in LDL Receptor Knockout (LDLRKO) Mice, Diabetes. 2016; 65(6): 1565-1576. doi: 10.2337/db15-0917.
- T Duan et al. Broccoli-Derived Exosome-like Nanoparticles Alleviate Loperamide-Induced Constipation, in Correlation with Regulation on Gut Microbiota and Tryptophan Metabolism, J Agric Food Chem. 2023; 71(44): 16568-16580. doi: 10.1021/acs.jafc.3c04150.
- Turmeric and Its Major Compound Curcumin on Health: Bioactive Effects and Safety Profiles for Food, Pharmaceutical, Biotechnological and Medicinal Applications - PMC. Accessed. 2023. [Online]. Available: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7522354/.80.
- S Fuloria et al. A Comprehensive Review on the Therapeutic Potential of Curcuma longa Linn. in Relation to its Major Active Constituent Curcumin, Front Pharmacol. 2022; 13: 820806. doi: 10.3389/fphar.2022.820806.
- Y He, Y Yue, X Zheng, K Zhang, S Chen, and Z Du, Curcumin, inflammation, and chronic diseases: how are they linked?, Molecules. 2015; 20(5): 9183-9213. doi: 10.3390/molecules20059183.
- L Vecchi Brumatti, A Marcuzzi, PM Tricarico, V Zanin, M Girardelli, and AM Bianco, Curcumin and inflammatory bowel disease: potential and limits of innovative treatments, Molecules. 2014; 19(12): 21127-21153, Dec., doi: 10.3390/molecules191221127.
- C Liu et al. Oral administration of turmeric-derived exosome-like nanovesicles with anti-inflammatory and pro-resolving bioactions for murine colitis therapy, Journal of Nanobiotechnology. 2022; 20(1): 206. doi: 10.1186/s12951-022-01421-w.
- M Zu et al.“Green” nanotherapeutics from tea leaves for orally targeted prevention and alleviation of colon diseases, Biomaterials. 2021; 279: 121178. doi: 10.1016/j.biomaterials.2021.121178.
- D Chen, G Chen, C Chen, X Zeng, and H.Ye, Prebiotics effects in vitro of polysaccharides from tea flowers on gut microbiota of healthy persons and patients with inflammatory bowel disease, Int J Biol Macromol. 2020; 158: 968-976. doi: 10.1016/j.ijbiomac.2020.04.248.
- Selenium-containing tea polysaccharides ameliorate DSSinduced ulcerative colitis via enhancing the intestinal barrier and regulating the gut microbiota - ScienceDirect. Accessed. 2023. [Online]. Available: https://www.sciencedirect.com/science/article/abs/pii/S0141813022007255
- C Kürbitz et al. Epicatechin gallate and catechin gallate are superior to epigallocatechin gallate in growth suppression and antiinflammatory activities in pancreatic tumor cells, Cancer Sci. 2011; 102(4): 728-734. doi: 10.1111/j.1349-7006.2011.01870.x.
- YS Chiou, Q Huang, CT Ho, YJ Wang, and MH Pan, Directly interact with Keap1 and LPS is involved in the anti-inflammatory mechanisms of (-)-epicatechin-3-gallate in LPS-induced macrophages and endotoxemia, Free Radic Biol Med. 2016; 94: 1-16. doi: 10.1016/j.freeradbiomed.2016.02.010.
- Molecules | Free Full-Text | Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Accessed. 2023. [Online]. Available: https://www.mdpi.com/1420-3049/22/3/484.
- MK Sriwastva et al. Exosome-like nanoparticles from Mulberry bark prevent DSS-induced colitis via the AhR/COPS8 pathway, EMBO Rep. 2022; 23(3): 53365. doi: 10.15252/embr.202153365.
- L Zhou, Y Jiang, Q Luo, LLi, and L Jia, Neddylation: a novel modulator of the tumor microenvironment, Mol Cancer. 2019; 18(1): 77. doi: 10.1186/s12943-019-0979-1.
- Cullin-RING ubiquitin E3 ligase regulation by the COP9 signalosome - PubMed. Accessed. 2023. [Online]. Available: https://pubmed.ncbi.nlm.nih.gov/27029275/.
- Y Luo and Y Song, Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities, Int J Mol Sci. 2021; 22(21): 11401. doi: 10.3390/ijms222111401.
- N Mookherjee, MA Anderson, HP Haagsman, and DJ Davidson, Antimicrobial host defence peptides: functions and clinical potential, Nat Rev Drug Discov. 2020; 19(5): 311-332. doi: 10.1038/s41573-019-0058-8.
- Chemical composition of Momordica charantia L. fruits | Journal of Agricultural and Food Chemistry. Accessed. 2023. [Online]. Available: https://pubs.acs.org/doi/abs/10.1021/jf00010a013.
- S Sur and RB Ray, Bitter Melon (Momordica Charantia), a Nutraceutical Approach for Cancer Prevention and Therapy, Cancers. 2020; 12(8): 8. doi: 10.3390/cancers12082064.
- PR Dandawate, D Subramaniam, SB Padhye, and S Anant, Bitter melon: a panacea for inflammation and cancer, Chinese Journal of Natural Medicines. 2016; 14(2): 81-100. doi: 10.1016/S1875-5364(16)60002-X.
- A Semiz, OO Acar, H Cetin, G Semiz, and A Sen, Suppression of inflammatory cytokines expression with bitter melon (Momordica charantia) in TNBS-instigated ulcerative colitis, Journal of Translational Internal Medicine. 2020; 8(3); 177-187. doi: 10.2478/jtim-2020-0027.
- F Wang, M Yuan, C Shao, N Ji, H Zhang, and C Li, Momordica charantia-Derived Extracellular Vesicles Provide Antioxidant Protection in Ulcerative Colitis, Molecules. 2023; 28(17): 17, Jan. doi: 10.3390/molecules28176182.