
Journal of Gastroenterology Research and Practice
Research Article - Open Access, Volume 3
Effects of hydroxychloroquine on intestinal mucosal barrier and gut microbiota in inflammatory-bowel disease of mice
Pan Huang; Yan Zhou; Zhipeng Wu; Siyu Wang; Caifang Ren; Genbao Shao; Zhengrong Zhou*
School of Medicine, Jiangsu University, Zhenjiang 212013, PR China.
*Corresponding Author : Zhengrong Zhou
School of Medicine, Jiangsu University, 301 Xuefu
Road, Zhenjiang 212013, China.
Email: zrzhou@ujs.edu.cn
Received : Apr 03, 2023
Accepted : May 05, 2023
Published : May 12, 2023
Archived : www.jjgastro.com
Copyright : © Zhou Z (2023).
Abstract
Hydroxychloroquine (HCQ) effect on gut microbiota and intestinal mucosal barrier in Inflammatory-Bowel Disease (IBD) was evaluated in this work. Here, we established an IBD model before treating mice with HCQ. After HCQ treatment, the body weight, colon length and fecal occult blood of mice did not change significantly. The structure and morphology of intestinal mucosa recovered, the expression of tight junction proteins (E-cadherin and Occludin) increased, the permeability of intestinal mucosal barrier decreased, the serum IL-10 increased, and the TNF-α decreased. Also, HCQ increased the abundance of Euryarchaeota_unclassified, Lactobacillus_murinus and Clostridium_fusiformis, but decreased the abundance of Oscillibacter, uncultured_Odoribacter, Bacterioidetes_unclassified, Para-bacteroides_distasonis and Muribaculum. These results demonstrate a positive impact of HCQ on the damage caused by IBD.
Keywords: IBD; HCQ; Intestinal injury; Gut microbiota; Intestinal mucosal barrier.
Citation: Huang P, Zhou Y, Wu Z, Wang S, Zhou Z, et al. Effects of hydroxychloroquine on intestinal mucosal barrier and gut microbiota in inflammatory-bowel disease of mice. J Gastroenterol Res Pract. 2023; 3(4): 1142.
Introduction
A recurrent and long-lived inflammation disorder of gastro-intestinal tract is termed as Inflammatory-Bowel Disease (IBD), which is mediated by imbalance of immune system of Intestinal Mucosa (IM) [1]. Treatment cost of IBD is estimated to be high in the midst of more than one million of American natives and two million five hundred thousand Europeans having the disease. In recent times, the disease has become a world-wide health problem as its frequency is increasing globally more especially in industrial nations in Middle East, Asia and South America [2]. Two subclasses of IBD have been established, namely Crohn disease and ulcerative colitis. Existing theory of IBD causation suggests interplays among risk factors such as immune system, microbes and environment in hosts who are genetically vulnerable [1]. Pathologically, alterations in tissues of intestines and dysfunction of enteric barrier of epithelium is induced by IBD. Despite incomplete understanding of IBD etiology, the relationship of the disease development with microbiota of the intestines is well established [3]. Therefore, there is an unmet need for discovering IBD treatment by identifying effective drugs to repair intestinal injury and regulate gut microbiota.
Lysosomotropic drugs like hydroxychloroquine (HCQ) are alkaline in nature with increased lysosomal accumulation, wherein they suppress significant biological processes through pH increment [4]. Presently, HCQ is considered as one of the important drugs for treating rheumatic disorder in the clinics, particularly rheumatoid arthritis and systemic-lupus erythematosus [4]. Through earlier works, health benefits of HCQ have been known to include anti-inflammation and immunomodulation [5]. In particular, HCQ reduced proinflammatory cytokines levels that were generated by peripheral mononuclear blood cells by inhibiting toll like receptor (TLR) signaling in endosomes, wherein the cytokines included interleukin-1 (IL-1), IL-2, IL-6, Interferon-Gamma (IFN-γ) and Tumor-Necrosis Factor-Alpha (TNF-α) [6]. Consequently, HCQ decreased deviant immune response activation which is mediated by TLR signaling, amid reduction of inflammatory indications in individuals with rheumatic disease [7]. Moreover, recent works have described HCQ effect on pathways of metabolism, platelets and neoplasm [8].
Materials and methods
Animal experiment protocol
Jiangsu University’s ethics committee gave approval (SCXK2018-0012) to the protocol of this experiment prior to commencement. The Center of Animal Laboratory at Jiangsu University (Jiangsu, China) supplied female C57BL/6J mice (10 weeks old), wherein they were acclimatized to laboratory environment without pathogens at 22 ± 3OC temperature and 40-60% humidity. We allocated the mice into 3 groups at random with each group comprising of 7 mice, wherein they were studied for a period of 3 weeks. While mice in CON group were given demineralized water, their counterparts in DSS group received 3% DSS in their drinking water while those in HCQ group were supplemented with HCQ (5mg/mL) in their drinking water.
All the mice were given standard food they were later euthanized within 3 weeks. We sampled blood from their retro-orbit, wherein we separated the serum from clotted blood through 15 min of centrifugation at 3000 × g and 4OC. Later on, anesthesia of mice was carried out with ether before they euthanized via ligation of cervices. Afterwards, we divided tissues of colon before storage in a freezer (-80OC). Prior to freezing in -80OC for 16S rDNA sequencing, we removed the intestinal feces and stored in liquefied nitrogen gas.
Enzyme-linked immunosorbent assay (ELISA) for cytokines
The concentrations of cytokines, including IL-4 (QZ-10258), IL-6 (QZ-10260), IL-10 (QZ-10235), and TNF-α (QZ-10225), were measured in the serum using commercial ELISA kits (all purchased from JiuBang Biotech, Fujian, China) according to the manufacturer’s protocols.
Analysis of morphology and immunohistochemical (IHC)
Phosphate Buffered Solution (PBS) was used to wash paraformaldehyde (4%)-fixed tissues before dehydration with ethanol (70%, 95% and 100%) and anhydrous alcohol. Later, we sectioned (5 μm thick) the colon tissues after they have been made transparent by xylene, coupled with paraffin embedment. Later, staining of the sections was accomplished with Alcian Blue (Leagene, DG0041) and hematoxylin-eosin (H&E) (SolarBio, DSS120).
We carried out IHC with the under listed antibodies antiClaudin-1 (1:200, 28674-1-AP, Proteintech, USA), anti-Occludin (1:200, EPR20992, Abcam, UK) and anti-E-Cadherin (1:200, 24E10, CST, MA, USA) and Rabbit two-step detection kit (PV-9001, ZSGB-BIO, Beijing, China). All-night incubation of paraffinized sections with primary antibody was carried out at 4OC after which we incubated for 20 minutes with accompanying secondary antibody at 37OC. Staining of the nuclei with hematoxylin and subsequently with DAB was carried out in accordance with manufacturer’s protocol. Optical microscope was employed to examine the sectioned tissues.
Fluorometric assay of FITC -dextran 4
Mice were fasted for 12 hours before the experiment. Mice in each group were given FITC-dextran 4 500 mg/kg by gavage. After 2 hours, the mice were anesthetized with chloral hydrate for imaging in vivo. After four hours, mice were sacrificed by CO2 asphyxiation. Blood was collected by cardiac puncture and serum were separated by centrifugation at 3,000 rpm, for 15 minutes at 4°C. FITC-dextran 4 in serum was determined at the excitation wavelength of 493 nm and the emission wavelength of 518.5 nm.
Analysis of gut microbiota
Based on protocol of the manufacturer, we performed sequencing of samples on the platform Illumina NovaSeq, which was derived from LC-Bio. In analyzing species diversity complexness, we applied alpha diversity via five indicants, namely Observed species, Goods coverage, Chao1, Simpson and Shannon. The indicants employed in this work were computed via QIIME2. In particular, calculation of beta diversity was accomplished with QIIME2, while plotting of data was performed with R-package. We calibrated the sequence with Blast, wherein SILVA database was used to annotate each illustrative sequence. Imple- mentation of other diagrams was done with R-package (V3.5.2).
Statistical analysis
We presented the data with means and standard error of mean (SEM). Assessment of groups’ differences was accomplished with unpaired Student t-test. Evaluation of more than 2 data sets was done with Kruskal-Wallis test, while analysis of correlations was carried out with Spearman’s ρ (rho). Graph-pad prism 8 version (Graph-pad Software) was employed to analyze the data, wherein in terms of statistics we accepted p<0.05 as significance level.
Results
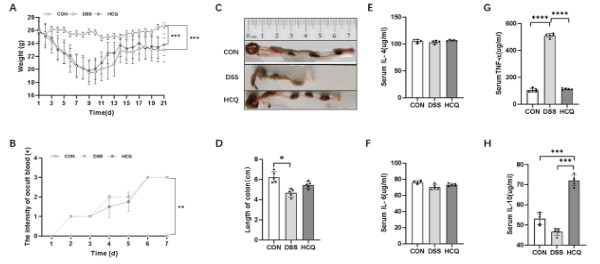
Effect of HCQ on body weight of mice
We measured the body weight of mice daily and found significant weight loss in the DSS group compared with the CON group (p<0.001, Figure 1A). However, the weight of mice in HCQ group was significantly lower than that in CON group (p<0.001, Figure 1A), but there was no significant difference between HCQ group and DSS group.
Effect of HCQ on the length of colon and the intensity of occult blood
Shorter length of colon was markedly seen mice of DSS group compared with CON group (p<0.05, Figure 1C), while in DSS group, the mice had serious fecal occult blood compared to those in CON group (p<0.01, Figure 1B). We observed no significant changes in colon length and occult blood intensity in HCQ group compared with the DSS group (Figure 1B and 1D).
Effect of HCQ on serum inflammatory cytokines concentrations
There was no significant difference in serum IL-4 and IL-6 concentrations among the three groups (Figure 1E and 1F). However, we found marked increased serum TNF-α concentrations in mice of DSS group than those in CON and HCQ groups (p<0.001) (Figure 1G). Of note, lower serum IL-10 concentration was seen in mice allocated to CON and DSS groups compared to those of HCQ group (p<0.001, Figure 1H).
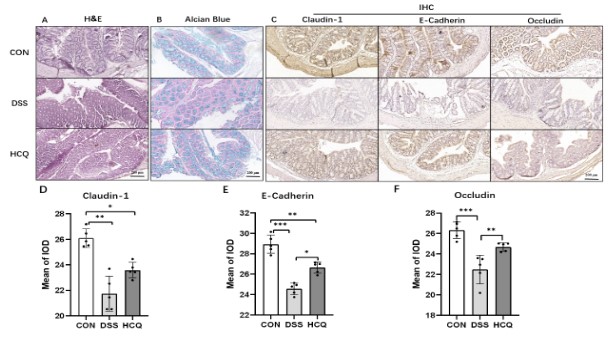
HCQ effect on morphology of intestinal mucosa
Figure 2A presented colon structure of mice in CON group, wherein it was intact with clear mucosa and orderly arranged glands, coupled with no obvious inflammatory cell infiltration. The structure of colon in DSS group was severely damaged with unclear structure of intestinal wall and disorderly arranged glands compared to those categorized under CON group. Also, in the aforementioned mice, we observed plentiful neutrophils and lymphoplasma cell infiltrations in mucosa and submucosa, wherein the crypt structure was break with loss of goblet cells. After treatment of HCQ, mice in HCQ group showed significant improvement compared to DSS group. The inflammatory cells in intestinal mucosa were decreased, while the structure of intestinal wall became clear. Meanwhile, the glands were arranged orderly, with the recovery of crypt shape and increase in goblet cells. Furthermore, we discovered through staining of sectioned tissues with Alcian Blue that thickness of mucus decreased substantially in animals of DSS group compared to those allocated to the CON group. Noticeably, those treated with HCQ demonstrated increasingly produced mucus compared to DSS group (Figure 2B).
Detection of localization and expressions of Claudin-1, E-Cadherin and Occludin were carried out in epithelium of enteric mucosa to explore the mechanistic action of HCQ (Figure 2C). Observably, the proteins were mainly expressed in cytosol of epithelium cells’ cytoplasm. Comparatively, we observed marked reduced expression of the aforesaid proteins in DSS group than in CON group. The expression of E-cadherin and Occludin increased significantly after HCQ treatment (Figure 2D-F).
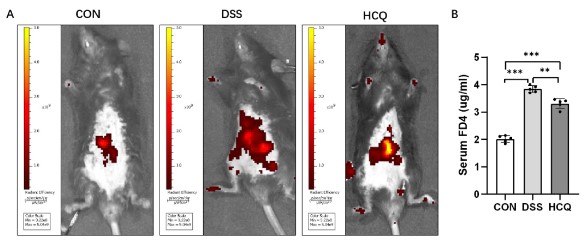
Effect of HCQ on intestinal mucosal permeability in mice
Two hours after instillation of FITC-dextran 4 in mice, the leakage of FITC-dextran 4 was observed by in vivo imaging, and the colonic mucosal permeability was significantly increased in the DSS group compared with the CON group. Compared with DSS group, HCQ group had significantly less leakage area and increased permeability (Figure 3A). Compared with CON group, the serum FD4 content in DSS group and HCQ group was significantly increased. Compared with DSS group, serum FD4 was significantly decreased in HCQ group (Figure 3B).
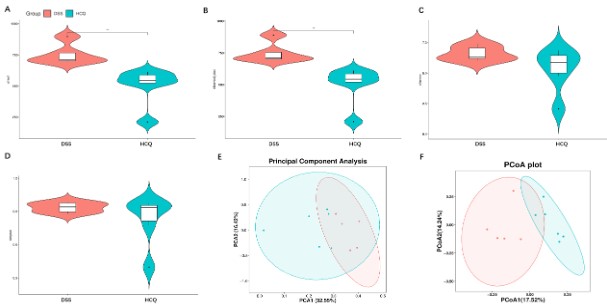
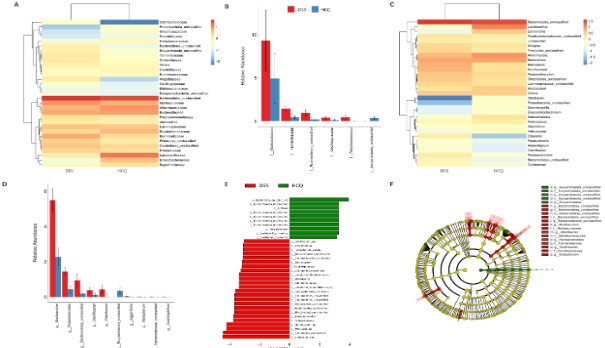
Effects of HCQ on gut microbiota diversity
We measured several indexes of alpha diversity in order to analyze alterations in gut microbiota diversity of DSS group and that of HCQ group. As the shown by charts, the Chao1 and Observed_otus index represented the number of species contained in a community (Figure 4A-B), while indexes of Simpson and Shannon ponder diversity and uncertainty (Figure 4C-D). After the treatment of the mice with HCQ, it was indicated that Chao1 and the Observed otus index were decreased significantly, while insignificant differences in indexes of Simpson and Shannon were observed (Figure 4A-D).
Both analyses of principal component and principal coordinates belong to beta diversity, wherein they represented the differences in species between different communities. We observed that HCQ treatment increased difference in intestinal species (Figure 4E-F).
Effect of HCQ on gut microbiota species abundance
We also compared the differences in species abundance of gut microbiota in DSS group with those allocated to HCQ group. With regards to family level, we observed a substantial increased (p<0.05 for all) abundance of Muribaculceae, Tannerellaceae, Bacterioidetes_unclassified, Oscillospiraceae, Odoribacteraceae in DSS group comparable to HCQ group. Meanwhile, we found a lowered abundance of Euryarchaeota_unclassified (P<0.05) in DSS group compared to HCQ group (Figure 5A-B). Comparison at genus level, showed that Muribaculum, Parabacteroides, Bacterioidetes_unclassified, Oscillibacter, Odoribacter abundances markedly increased (p<0.05 for all) in DSS group compared to HCQ group. However, Euryarchaeota_unclassified (p<0.05 for all) abundance was lower in DSS group compared to HCQ group (Figure 5C-D).
The LDA effect size (LEfse) mainly aims to compare two or more groups, wherein it is used to identify species that significantly differ in terms of abundance within distinct groups (biomarker) (Figure 5E-F). Comparison at species level, showed that Euryarchaeota_unclassified, Lactobacillus_murinus and Clostridium_fusiformis abundances increased in HCQ group compared to DSS group. However, Oscillibacter, uncultured_Odoribacter, Bacterioidetes_unclassified Parabacteroides_distasonis and Muribaculum abundance were lower in HCQ group compared to DSS group (Figure 5E).
Discussion
As a well-known anti-inflammatory drug, HCQ is described to inhibit inflammatory reaction through the following means [6]. (i) Activated TLR4 induced generation of ROS and proinflammatory mediators in macrophages is inhibited by HCQ [10]. (ii) Activated NLRP3 inflammasome in-vivo and in-vitro is suppressed by HCQ, amid the innate immune system cytoplasmic component influencing initial response to inflammation [11]. (iii) The assembling of NADPH-oxidase (NOX) in the endosomes is interfered by HCQ wherein the former process occurs in lots of prothrombotic and inflammatory pathways [12]. In this regard, we detected serum concentrations of inflammatory cytokines in IBD mice. The results are consistent with earlier works [13,14]. When pathogenic microorganisms invade the body, they activate themselves by recognizing receptors on the surface of antigens, and by enhancing phagocytosis, secrete various pro-inflammatory cytokines and anti-inflammatory factors to clear pathogens, so as to clear pathogens and control the disease process inflammatory cytokines, such as TNF-α, IL-6 and IL-10 are the main factors involved in the regulation of immune response in IBD. The expression and regulation of these cytokines interact and coordinate with each other [15,16]. An increased serum IL-10 concentration and decreased serum TNF-α level indicate that HCQ treatment could reduce inflammatory reaction. Nevertheless, the results suggest that HCQ had no significant effect on the length of colon, body weight and intensity of fecal occult blood after the mice were treated with HCQ. Gastrointestinal bleeding and loss of body weight are common clinical symptoms of IBD [17], thereby relieving IBD patients from these symptoms is an important aspect of the disease treatment [18].
Inflammation and injury of the gastrointestinal epithelium can induce mucosal barrier dysfunction, leading to barrier defects, and increase the migration of FITC-dextran 4 molecules into the intestinal serosa layer, and then into the systemic circulation. After oral administration, FITC-dextran 4 molecules can cross the damaged intestinal epithelial cells and be quantified in the blood Serum FITC-dextran 4 concentration can be used as an index to measure the degree and severity of intestinal mucosal barrier dysfunction in paracellular permeability [19-21]. In this study, the effect of hydroxychloroquine treatment on colonic mucosal permeability in mice was studied by in vivo imaging tracer experiment and the serum fluorescence intensity of FITC-dextran 4 was measured. The results indicated that hydroxychloroquine treatment may improve the damage of intestinal mucosal barrier induced by DSS by reducing the colonic mucosal permeability.
Goblet cells generate intestinal mucus, which is an important element of intestinal epithelial protection, coupled with the secretion of gel forming mucins [22]. As shown in our results, after we supplemented HCQ to the mice, the proportion of goblet cells and intestinal mucus as well as expression of E-Cadherin increased in IBD mice, while the shape of colonic crypt and intestinal wall also recovered. This finding indicates that HCQ has the capacity to protect the structure and morphology of colon. Also, tight-junction proteins were expressed in endothelia and epithelia, wherein they form barriers to para-cellular channel which influence permeability of tight-junction [23]. Loss of tight-junction proteins can induce intestinal epithelial dysfunction [24].
Gut microbiota is linked to progress of chronic disorders, viz., some tumors, metabolic syndrome and obesity [25-27]. Nowa-days the exact pathological process of IBD is still undisclosed, wherein scientists have generally accepted pathogenesis of IBD as a deviant response of immune system, which is induced by alterations in microbiota of the intestines [28]. In particular, it is mostly and consistently observed that bacterial diversity of intestinal microbiota is decreased in IBD patients, coupled with increased Proteobacteria and decreased Firmicutes [29]. We sequenced samples to study alterations in intestinal microbiota of mice with IBD. The results revealed that HCQ significantly decreased diversity of intestinal microbiota and decreased abundance of Bacteroides, Muribaculum, Odoribacter and Parabacteroides_distasonis, but increased abundance of Lactobacillus_murinus, Clostridium_fusiformis and Euryarchaeota_unclassified. Especially species of Bacteroides are important pathogens in the clinics, which beneficially relate to the host when they are held in the intestines, but they can cause serious pathology upon existing gut, amid having the most antibiotic resistance mechanisms [30,31]. In a mouse model of glucan sodium-induced colitis, the relative abundance of Muribaculaceae was negatively correlated with proinflammatory cytokines and positively correlated with the expression level of tight junction proteins. Therefore, the bacterial species belonging to the Muribaculum seems to be important for maintaining the normal condition of the mouse gut [32]. Odoribacter not only plays an excellent role in reducing intestinal inflammation, but also promotes intestinal maturation [33]. Parabacteroides_distasonis was identified as a recurring bacterium in the feces of CD patients. The potential evidence that Parabacteroides_distasonis may contribute to the pathogenesis of IBD remains in conclusive. Parabacteroides_distasonis can produce catalase, which allows it to combat oxidative stress mediated by hydrogen peroxide, usually produced by inflammatory cells. However, these oxidants may be inflammatory triggers for CD. This is because catalase catabolizes Reactive Oxygen Species (ROS) produced by various bacteria, including Parabacteroides_distasonis, which may exacerbate inflammation [29]. Besides, researches have verified that Lactobacillus_murinus could reduce inflammatory reactions [34]. Lactobacillus_murinus has been reported to successfully colonize the rodent gut and possess protective properties against necrotizing enterocolitis [26,35]. However, there are few studies about Euryarchaeota_unclassifiedand Clostridium_fusiformis in gut diseases. In IBD medication, there is complex and bi-directional interactions between gut microbiota and drugs, wherein the latter can influence constituent of the gut microbiota, which in turn can influence drug response of individuals [36]. The interaction between HCQ and gut microbiota still need further study. Notwithstanding, these results are meaningful for further investigations involving fecal microbiota transplantation and probiotics.
Taken together, these results suggest that supplementation of HCQ to IBD mice could reduce the inflammatory reaction, which protected the structure and morphology of colon by increasing goblet cells’ number and secretion of mucins, thus maintaining permeability of intestines via strengthen of tight-junction proteins’ expression. Also, we discovered that HCQ could regulate abundance of some intestinal probiotics and pathogens. Hence, HCQ has shown positive effects on the health of IBD mice gut, which suggest our findings have given a new approach to treating the disease.
Declarations
Funding: We appreciate our funders for their support. In particular, we received funding support for this work from Jiangsu Province Graduate Student Scientific Research Innovation project (KYCX22_3612), Zhenjiang Key R&D Program Social Development Project (SH2022037).
Disclosure statement: No potential conflict of interest was reported by the author(s).
Author contributions: Zhengrong Zhou, Pan Huang and Genbao Shao conceived and designed the experiments. Yan Zhou and Caifang Ren conducted the experiments. Yan Zhou, Siyu Wang and Zhipeng Wu collected the samples and analyze the data. Yan Zhou, Siyu Wang and Zhipeng Wu drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
References
- Glassner KL, Abraham BP, Quigley EMM. The microbiome and inflammatory bowel disease. J Allergy Clin Immunol. 2020; 145: 16-27.
- Kaplan GG. The global burden of IBD: from 2015 to 2025. Nat Rev Gastroenterol Hepatol. 2015; 12: 720-727.
- Weingarden AR, Vaughn BP. Intestinal microbiota, fecal microbiota transplantation, and inflammatory bowel disease. Gut Microbes. 2017; 8: 238-252.
- Ponticelli C, Moroni G. Hydroxychloroquine in systemic lupus erythematosus (SLE). Expert Opin Drug Saf. 2017; 16: 411-419.
- Li R, Lin H, Ye Y, Xiao Y, Xu S, et al. Attenuation of antimalarial agent hydroxychloroquine on TNF-alpha-induced endothelial inflammation. Int Immunopharmacol. 2018; 63: 261-269.
- Yao J, Xie J, Xie B, Li Y, Jiang L, et al. Therapeutic effect of hydroxychloroquine on colorectal carcinogenesis in experimental murine colitis. Biochem Pharmacol. 2016; 115: 51-63.
- Eugenia Schroeder M, Russo S, Costa C, et al. Pro-inflammatory Ca(++)-activated K(+) channels are inhibited by hydroxychloroquine. Sci Rep. 2017; 7: 1892.
- Tang TT, Lv LL, Pan MM, Wen Y, Wang B, et al. Hydroxychloroquine attenuates renal ischemia/reperfusion injury by inhibiting cathepsin mediated NLRP3 inflammasome activation. Cell Death Dis. 2018; 9: 351.
- Richard SA, Kampo S, Hechavarria ME, Sackey M, Buunaaim ADB, et al. Elucidating the Pivotal Immunomodulatory and Anti-Inflammatory Potentials of Chloroquine and Hydroxychloroquine. J Immunol Res. 2020; 2020: 4582612.
- Zeidi M, Kim HJ, Werth VP. Increased Myeloid Dendritic Cells and TNF-alpha Expression Predicts Poor Response to Hydroxychloroquine in Cutaneous Lupus Erythematosus. J Invest Dermatol. 2019; 139: 324-332.
- Muller-Calleja N, Manukyan D, Canisius A, Strand D, Lackner KJ. Hydroxychloroquine inhibits proinflammatory signalling pathways by targeting endosomal NADPH oxidase. Ann Rheum Dis. 2017; 76: 891-897.
- Rainsford KD, Parke AL, Clifford-Rashotte M, Kean WF. Therapy and pharmacological properties of hydroxychloroquine and chloroquine in treatment of systemic lupus erythematosus, rheumatoid arthritis and related diseases. Inflammopharmacology. 2015; 23: 231-269.
- Hu J, Wang X, Chen X, Fang Y, Chen K, et al. Hydroxychloroquine attenuates neuroinflammation following traumatic brain injury by regulating the TLR4/NF-kappaB signaling pathway. J Neuroinflammation. 2022; 19: 71.
- Lei ZN, Wu ZX, Dong S, Yang DH, Zhang L, et al. Chloroquine and hydroxychloroquine in the treatment of malaria and repurposing in treating COVID-19. Pharmacol Ther. 2020; 216: 107672.
- Marafini I, Sedda S, Dinallo V, Monteleone G. Inflammatory cytokines: from discoveries to therapies in IBD. Expert opinion on biological therapy. Nov 2019; 19: 1207-1217.
- Leppkes M, Neurath MF. Cytokines in inflammatory bowel diseases - Update 2020. Pharmacological research. 2020; 158: 104835.
- Bai L, Li J, Li H, Song J, Zhou Y, et al. Renoprotective effects of artemisinin and hydroxychloroquine combination therapy on IgA nephropathy via suppressing NF-kappaB signaling and NLRP3 inflammasome activation by exosomes in rats. Biochem Pharmacol. 2019; 169: 113619.
- Sasson AN, Ananthakrishnan AN, Raman M. Diet in Treatment of Inflammatory Bowel Diseases. Clin Gastroenterol Hepatol. 2021; 19: 425-435 e3.
- Woting A, Blaut M. Small Intestinal Permeability and Gut-Transit Time Determined with Low and High Molecular Weight Fluorescein Isothiocyanate-Dextrans in C3H Mice. Nutrients. 2018; 10.
- Vuong CN, Mullenix GJ, Kidd MT, Bottje WG, Hargis BM, et al. Modified serum fluorescein isothiocyanate dextran (FITC-d) assay procedure to determine intestinal permeability in poultry fed diets high in natural or synthetic pigments. Poultry science. 2021; 100: 101138.
- Li BR, Wu J, Li HS, Jiang ZH, Zhou XM, et al. In Vitro and In Vivo Approaches to Determine Intestinal Epithelial Cell Permeability. Journal of visualized experiments : JoVE. 2018; 140.
- Giambo F, Teodoro M, Costa C, Fenga C. Toxicology and Microbiota: How Do Pesticides Influence Gut Microbiota? A Review. Int J Environ Res Public Health. 2021; 18.
- Günzel D, Yu AS. Claudins and the modulation of tight junction permeability. Physiol Rev. 2013; 93: 525-569.
- Huang P, Jiang A, Wang X, Zhou Y, Tang W, et al. NMN Maintains Intestinal Homeostasis by Regulating the Gut Microbiota. Frontiers in Nutrition. 2021; 8.
- Hills RD, Pontefract BA, Mishcon HR, Black CA, Sutton SC, et al. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients. 2019; 11.
- Shang Z, Li M, Zhang W, Cai S, Hu X, et al. Analysis of phenolic compounds in pickled chayote and their effects on antioxidant activities and cell protection. Food research international (Ottawa, Ont). 2022; 157: 111325.
- Li M, Bao X, Zhang X, Ren H, Cai S, et al. Exploring the phytochemicals and inhibitory effects against α-glucosidase and dipeptidyl peptidase-IV in Chinese pickled chili pepper: Insights into mechanisms by molecular docking analysis. Lwt. 2022; 162.
- Matsuoka K, Kanai T. The gut microbiota and inflammatory bowel disease. Semin Immunopathol. 2015; 37: 47-55.
- Ezeji JC, Sarikonda DK, Hopperton A, Erkkila HL, Cohen DE, et al. Parabacteroides distasonis: intriguing aerotolerant gut anaerobe with emerging antimicrobial resistance and pathogenic and probiotic roles in human health. Gut Microbes. 2021; 13: 1922241.
- Zafar H, Saier MH. Gut Bacteroides species in health and disease. Gut Microbes. 2021; 13: 1-20.
- Wexler HM. Bacteroides: the good, the bad, and the nitty-gritty. Clin Microbiol Rev. 2007; 20: 593-621.
- Yan S, Yang B, Zhao J, Zhao J, Stanton C, et al. A ropy exopolysaccharide producing strain Bifidobacterium longum subsp. longum YS108R alleviates DSS-induced colitis by maintenance of the mucosal barrier and gut microbiota modulation. Food & function. 2019; 10: 1595-1608.
- Lima SF, Gogokhia L, Viladomiu M, Chou L, Putzel G, et al. Transferable Immunoglobulin A-Coated Odoribacter splanchnicus in Responders to Fecal Microbiota Transplantation for Ulcerative Colitis Limits Colonic Inflammation. Gastroenterology. 2022; 162: 166-178.
- Wang K, Liao M, Zhou N, Bao L, Ma K, et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019; 26: 222-235 e5.
- Isani M, Bell BA, Delaplain PT, Bowling JD, Golden JM, et al. Lactobacillus murinus HF12 colonizes neonatal gut and protects rats from necrotizing enterocolitis. PLoS One. 2018; 13: e0196710.
- Weersma RK, Zhernakova A, Fu J. Interaction between drugs and the gut microbiome. Gut. 2020; 69: 1510-1519.